
- Регуляция эстрального цикла крыс
- Аппаратный способ определения стадий эстрального цикла у мелких лабораторных грызунов
- Резюме
- Введение
- Реактивные изменения поведения крыс в динамике эстрального цикла.
- Содержание
- Введение
- Глава 1. Обзор литературы
- 1.1 Морфофункциональные изменения женской репродуктивной сֺиֺсֺтֺеֺмֺы
- 1.2 Понятие эстрального цикла и его этапы
- 1.3 Эстральный цикл у крысы
- 1.4 Поведенческие эффекты женских половых гормонов.
- 1.5 Поведенческие особенности крыс линии WAG/Rij
- Глава 2. Материал и методы исследования
- 2.1 Объекты исследования
- 2.2 Методы исследования
- 2.2.1 Определение стадий эстрального цикла самок крыс методом влагалищных мазков
- 2.2.2 Тест «Открытое поле»
- 2.2.3 Тест «Приподнятый крестообразный лабиринт»
- 2.2.4 Тест «Вынужденное плавание»
Регуляция эстрального цикла крыс
Эстральный цикл (от новолат. oestrus — течка), совокупность регулярно повторяющихся изменений половой системы у самок млекопитающих. Циклическая функция яичников, связанная с происходящими в них ритмическими процессами (развитие фолликулов, овуляция и образование жёлтого тела), гормональным путём определяет синхронные характерные изменения во всём организме самки, и особенно в половом аппарате (яйцеводы, матка и влагалище). Каждой фазе эстрального цикла соответствуют определённые изменения слизистой оболочки влагалища и клеточного состава влагалищного содержимого [65].
Эстральный цикл длится у крыс 4-5 суток. Весь эстральный цикл у грызунов условно разбивается на следующие стадии: диэструс (стадия покоя, или межтечки); проэтрус (стадия подготовки к течке, или предтечки); эструс (течка); метаэструс (стадия послетечки). Продолжительность эстрального цикла может меняться под действием различных факторов (стрес, нагрузки и пр.). У животных с эстральным циклом копуляция и оплодотворение могут происходить только во время овуляции.
Наиболее характерные циклические изменения у женских особей репродуктивного возраста проявляются в органах половой сферы — яичниках, матке и влагалище у млекопитающих. При этом периодические изменения в гонадах (овариальные циклы) являются ведущими по отношению к изменениям, возникающим в половом тракте.
Эстральные циклы протекают во влагалище и отражают циклические изменения вагинального эпителия, физиологически зависимые от эндокринных функций яичников [72].
1) Стадия проэструса продолжается около 12 часов, отражает наиболее высокий уровень секреции эстрогенов созревающими фолликулами яичника и характеризуется гипертрофией и гиперплазией эпителиальных клеток влагалища, а также последующим отторжением клеток, секретирующих слизь. В это время во влагалищном мазке обнаруживаются нормальные эпителиальные клетки [52].
2) Стадия эструса — завершение процессов активации вагинальных тканей эстрогенами и непосредственно следует за пиком выброса этих гормонов. Длительность стадии 27 часов. В этот период гипертрофированные и гиперплазированные клетки вагинального эпителия расслаиваются (стратифицируются) и образуют наружный чешуйчатый слой. В клетках чешуйчатого слоя откладывается кератин и ороговевшие клетки отторгаются. Это и есть собственно течка, отражающая наибольшую активность данной части полового тракта. В это время во влагалищном мазке обнаруживаются в основном ороговевшие клетки. Стадии эструса и конца проэструса сочетаются с активизацией полового поведения (состояние половой охоты) и по времени примерно совпадают с овуляцией, поэтому спаривание животных происходит нормально только в начале этой стадии (рис. 1.3 А) [52].
3) Стадия метаэструса характеризуется резким снижением активности вагинального эпителия, некробиотическими процессами и отторжением базального слоя клеток. Стадия метаэструса длится около 6 часов. На данной стадии в вагинальном мазке обнаруживаются ороговевшие и эпителиальные клетки, и множество лейкоцитов. Метаэструс — следствие снижения концентрации эстрогенов в крови. По времени эта стадия обычно совпадает с формированием желтых тел и началом секреции ими прогестинов [62].
4) Стадия диэструса — стадия покоя вагинального эпителия, составляет около половины всей продолжительности эстрального цикла. Полиферативные процессы в диэструсе заторможены, ороговения эпителиальных клеток не происходит, и они секретируют в полость влагалища слизь. Последняя, наряду с лейкоцитами, обнаруживается в вагинальном мазке. Состояние диэструса обусловлено в значительной степени низким уровнем эстрогенов в организме. Однако секреция прогестинов желтыми телами в этот период цикла также вносит свой вклад в вагинальные процессы. Гестагены тормозят разрастание, стратификацию и ороговение вагинального эпителия и стимулируют секрецию слизи поверхностным слоем клеток. Секретируемая в просвет влагалища слизь препятствует продвижению сперматозоида в матку. В отношении клеток влагалища прогестины являются антагонистами эстрогенов (рис. 1.3 Б) [58].
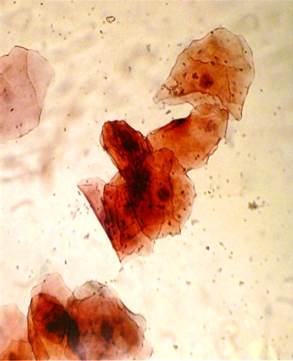

Рис. 1.3. Вагинальные мазки. (А). Эструс. (Б) Диэструс [95].
Таким образом, эстральный цикл имеет значение для репродуктивной активности животных. Диэструс покой вагинального эпителия, обусловленный низким уровнем эстрогенов в организме; на него падает около половины продолжительности эстрального цикла. Лизис жёлтого тела и переход яичника в фолликулярную фазу овариального цикла служат сигналом к началу нового эстрального цикла и переходу от диэструса в проэструс.
Стадия диэструса интересна тем, что в этот период исчезают признаки течки и полового возбуждения. Во влагалищных мазках преобладают клетки лейкоцитов. Диэструс является самой активной частью лютеиновой фазы полового цикла, которая начинается сразу же после овуляции. Потому в данном исследовании изучалась роль задних отделов миндалевидного комплекса в организации поведения самок крыс именно на стадии диэструса.
Источник
Аппаратный способ определения стадий эстрального цикла у мелких лабораторных грызунов
И.В. Белозерцева(1), кандидат биологических наук, зав. отделом психофармакологии, зав. лабораторией экспериментальных доклинических исследований с виварием; Д.Д. Меркулов(2), доктор инженерных наук (Dr. sc. ing.), директор проектов ELMI Ltd. О.Е. Вилитис(3), доктор физических наук (Dr. phys.), Заслуженный деятель науки (State Emiritus Scientist); Б.В. Скрябин(4), MD, PhD, руководитель «Transgenic animal and genetic engineering Models (TRAM) facility» 1-Институт фармакологии им. А.В. Вальдмана, ФГБОУ ВО «Первый Санкт-Петербургский государственный медицинский университет им. И.П. Павлова» Минздрава России, 197022, Россия, Санкт-Петербург, ул. Льва Толстого, 6 –8; 2-ELMI Ltd., LV-1006, Латвия, Рига, ул. Криву, 11; 3-Институт физики твердого тела, Латвийский университет, Лаборатория органических материалов, LV-1063, Латвия, Рига, ул. Кенгарага 8; 4-Медицинский факультет университета г. Мюнстер, Германия (TRAM, Faculty of Medicine, WWU Muenster), D-48149 Muenster, Germany, Von Esmarch St. 56 E-mail: belozertseva@gmail.com
Резюме
Резюме. На самках мышей и крыс выполнена апробация нового инструментального метода/устройства для определения стадий эстрального цикла у мелких грызунов. Циклические изменения активного сопротивления (RS) слоя эпителиальных клеток слизистой оболочки влагалища измеряли с помощью нового детектора MED-PRO 100 (Elmi Ltd., Latvia). Активное сопротивление измеряли на частоте 1 кГц, вставляя зонд детектора (l = 14 мм, d = 1,82 мм; диапазон измерения – 0–50 кОм) во влагалище животного на 1–2 с. Результаты измерений соотносили с цитологической картиной влагалищных мазков, классического способа определения стадий эстрального цикла. Было установлено, что новый метод дает точное определение стадии в «пиковых» значениях и его целесообразно использовать, поскольку он нетравматичен для животного и не так трудоемок, как классический цитологический метод. Построенные кривые активного сопротивления для нескольких эстральных циклов были схожи (особенно у крыс), а изменения активного сопротивления соответствовали стадиям, определяемым по цитологической картине мазков. Однако максимальные значения активного сопротивления соответствовали разным стадиям цикла у мышей и крыс. У крыс активное сопротивление было значимо больше в проэструсе (>5 кОм) по сравнению с другими стадиями цикла и в метэструсе по сравнению с эструсом и диэструсом. У мышей максимальному значению активного сопротивления соответствовала стадия эструса. Мы пришли к выводу, что использование устройства MED-PRO 100 обеспечивает точный подход к оценке эстрального цикла у крыс и мышей. Это может быть использовано для определения оптимального времени скрещивания животных; гормональной суперовуляции или технологий искусственного оплодотворения, а также для оптимизации поведенческих, фармакологических и токсикологических исследований, выполняемых на самках лабораторных грызунов.
Введение
Проведение фармакологических [1, 2] и токсикологических [3] исследований на самках лабораторных грызунов; выбор оптимального времени скрещивания для получения беременных (или псевдобеременных) самок при создании трансгенных животных или искусственном оплодотворении [3, 5, 6, 7], поддержания колоний лабораторных животных, особенно линий с низкой фертильностью [8], а также вскрытие для морфологических исследований [3], требуют определения стадии эстрального цикла. Учет данного фактора может оптимизировать выполнение «профильных» экспериментов и повысить результативность воспроизводства лабораторных животных. Начиная с 20-х годов ХХ века [9, 10] используется метод оценки цитологической картины смыва или мазка из влагалища самки («золотой стандарт») для определения стадии эстрального цикла, в котором она находится. Данная процедура выполняется в несколько этапов, требующих определенных временных затрат, а также наличие микроскопа и ряда расходных материалов – пипеток, предметных стекол, красителей и пр. При этом способы окрашивания мазков и субъективность анализа готовых препаратов предварительно обученным персоналом также могут влиять на конечные результаты. Кроме прочего было показано, что многократные ежедневные смывы и взятие мазков с помощью тампонов могут нарушать протекание циклов [11] и даже вызывать ороговение клеток эпителия влагалища у овариэктомированных самок крыс [12]. В связи с этим, поиск быстрого и надежного аппаратного способа определения стадий полового цикла у самок лабораторных животных представляется весьма актуальной задачей.
Разработки детекторов для определения времени овуляции по электрическому сопротивлению стенок влагалища у сельскохозяйственных видов животных начались довольно давно. Еще в 1961 г. в СССР было оформлено авторское свидетельство «Способ определения оптимального времени осеменения коров» [13], а чуть позже был получен патент №178602 на прибор для измерения электрического сопротивления стенок влагалища у коров [14]. Упоминание о детекторах стадий полового цикла у лабораторных грызунов – крыс [15, 16, 17] и морских свинок [18] начали появляться с конца 1970-х гг. Однако до настоящего времени среди доступных коммерческих детекторов не существует надежного аппарата для мышей, о чем некоторые производители данного оборудования открыто предупреждают на своем сайте (например, для Model МК-12 (Muromachi Kikai Co., Ltd., Япония) – https://muromachi.com/en/archives/english/2108/).
В 2015 г. в компании Elmi (Латвия) была начата разработка детектора MED-PRO (рис. 1), ориентированного в первую очередь на определение стадий эстраль-ного цикла у мышей. В настоящее время эта разработка защищена патентом LV15278 [19].

В существующих детекторах эстрального цикла грызунов электрическое сопротивление слизистой влагалища определяется измерением электрического импеданса (Z), который представляет собой геометрическую сумму активного сопротивления (АС) электрической цепи RS и реактивного сопротивления XС :
RS является величиной, обратной величине электропроводности ткани, которая мало зависит от частоты переменного тока, в то время как реактивная составляющая XС импеданса зависит от частоты электрического тока.
При измерениях на переменном токе Z зависит от наличия границ раздела в системе, на которых может происходить накопление зарядов – поляризация. Свойства границ раздела (в биологическом объекте это главным образом различные мембраны клеток) могут быть описаны, если учесть емкость С, сопротивление которой XС зависит от частоты, на которой производится измерение:
где ω = 2π f – круговая частота, f – частота в Гц.
Измерение электропроводимости биологических систем на постоянном токе крайне затруднено из-за высокой степени поляризации на поверхностях мембран клеток и электродов зонда, поэтому для измерений применяют переменный ток.
Для существенного повышения точности измерений в новом детекторе эстрального цикла MED-PRO 100 применен метод синхронного детектирования [19], который обеспечивает селективное измерение только компоненты активной части сопротивления RS, устраняя, таким образом, влияние C на результат измерения.
В отличие от массивных зондов у ныне существующих детекторов эстрального цикла, требующих времени на температурную стабилизацию показателя, созданный в Elmi детектор MED-PRO 100 имеет зонд миниатюрных размеров (рис. 1, справа). Он имеет два электрода (∅ = 1,82 мм, шириной по 1,2 мм,
материал – AISI316, зазор между электродами – 1,2 мм) с малой теплоемкостью способствует быстрому ( 15 мес) и полностью сглаживаются к 20 месячному возрасту.
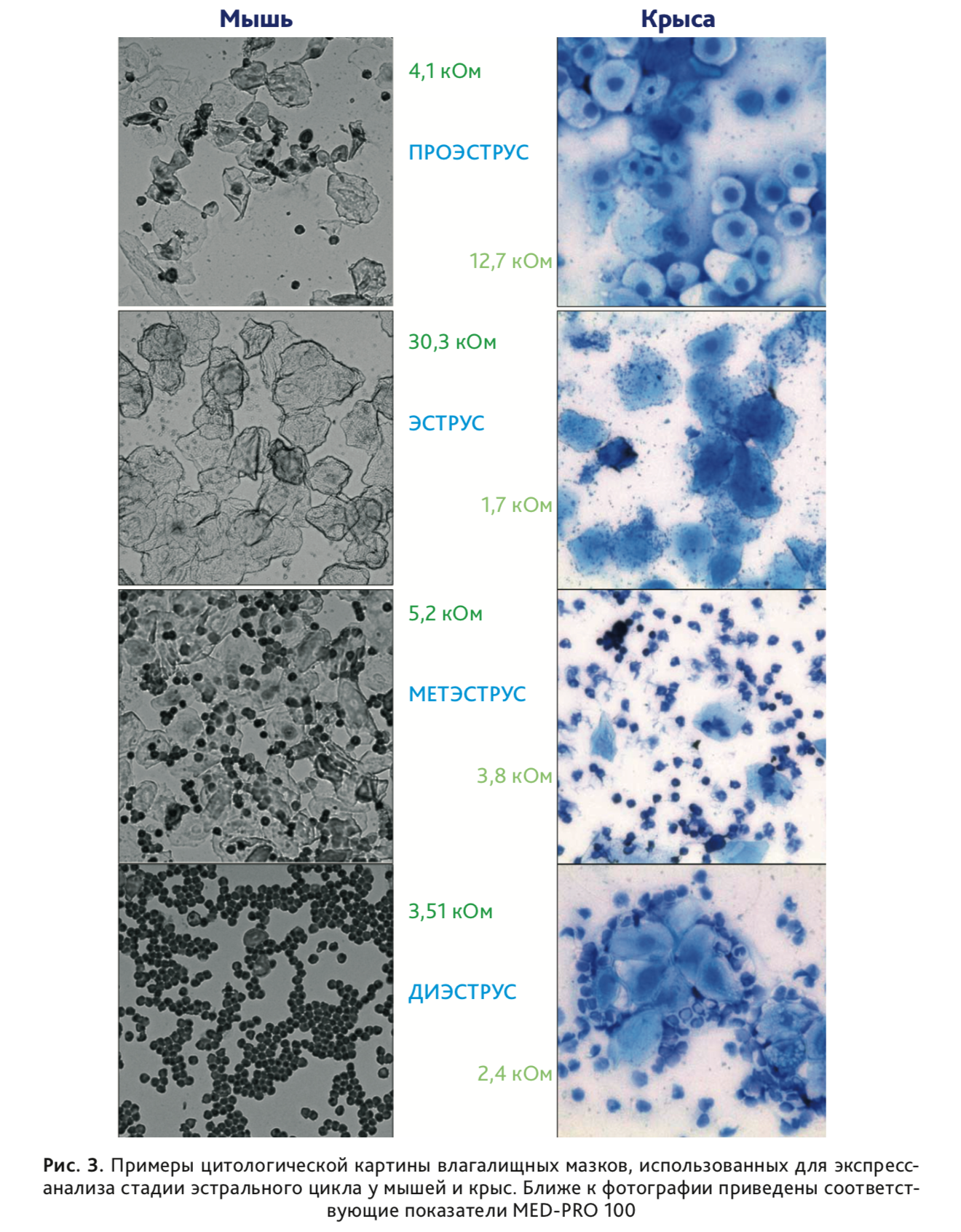
В отличие от самок мышей, у крыс стока Wistar в возрасте 3 мес, содержащихся в группах однопометников, наблюдали регулярные циклы колебания АС стенок влагалища. Замеры производили ежедневно за 1-2 часа до наступления темной фазы суточного цикла (12 ч свет /12 ч темнота). У 72% самок (n=61) длительность цикла составляла 4 дня (рис. 4). Усредненные по 5 циклам показатели АС стенки влагалища (n=44) представлены на рис. 5. Однофакторный дисперсионный анализ (на рангах) с повторными измерениями показал значимые различия АС стенки влагалища у крыс, находящихся в разных фазах эстрального цикла (Fr=476,7, df=3, p
Источник
Реактивные изменения поведения крыс в динамике эстрального цикла.
Страницы: 1 2
Содержание
- Введение
- Глава 1. Обзор литературы
- 1.1 Морфофункциональные изменения женской репродуктивной системы
- 1.2 Понятие эстрального цикла и его этапы
- 1.3 Эстральный цикл у крыс
- 1.4 Поведенческие эффекты женских половых гормонов
- 1.5 Поведенческие особенности крыс линии WAG/Rij
- Глава 2. Материал и методы исследования
- 2.1 Объекты исследования
- 2.2 Методы исследования
- Глава 3. Результаты исследования и их обсуждение
- 3.1 Изучение эстрального цикла методом влагалищных мазков
- 3.2 Исследование уровня тревожности крыс в зависимости от эстрального цикла
- 3.3 Оценка уровня депрессивности самок крыс в зависимости от эстрального цикла
- 3.4 Оценка когнитивного поведения самок крыс в зависимости от эстрального цикла
- Выводы
- Список использованной литературы
Введение
В последние годы поведенческие особенности животных привлекают все внимание исследователей и ученых самых различных профессий.Институты физиологии и психологии исследуют поведение человека и животных с помощью разнообразных методов и подходов.Исследователям-зоологам при работе по систематике, исследований по ориентации и адаптации животных необходимо изучение поведения животного мира. При одомашнивании и содержании животноводами сельскохозяйственных животных необходима роль поведенческих особенностей. Кроме того, исследование поведения животных играет важную роль в развитии бионики[35].
Известно, что поведение — это физиологический процесс, представляющий собой адаптивную связь между организмом и окружающей средой. Ответная реакция организма изменяется в связи с изменением среды потому, что организмы взаимодействуют с живой или неживой окружающей средой. Поведение, как мы понимаем, это активные изменения или направленные действия, которые способствуют выживанию[18]. По литературным данным, эстрогены и прогестерон выполняют функцию, которая отвечает за контроль полового поведения.Как писал Бич, поведение самок значительно сложный процесс и его необходимо рассмотреть его в целом[41].
Теоретическую модель, предложенная Лешнером из обзора многочисленных исследований гормонов и агрессивного поведения, учитывает гормональный статус организма, влияние окружающей среды, развитие и свойства самого животного как объекта, вызывающего агрессивную реакцию [51].
Результаты, полученные в различных условиях на крысах разных линий (Wistar, Sprague-Dawley, Long-Evans, Fisher 344, WAG/Rij, SHR и др.), показывают существование определенной зависимости поведения от стадии эстрального цикла, что связывается с циклическими изменениями концентрации половых стероидов в плазме и в ткани мозга [11].
Поскольку, железы внутренней секреции и нервная система имеют общие функции коммуникации и координации внутри организма следует обратить внимание на взаимодействие половых гормонов и поведенческих особенностей. Гормоны, которые циркулируют в крови, часто рассматриваются как первичные мотивирующие факторы поведения животных, а измененный их уровень внутренними и внешними факторами отражается на особенностях поведения. Работы, посвященные ориентировочно-исследовательской деятельности занимают основное место в психофизиологических исследованиях. Однако воздействие гормональных факторов на высшие функции головного мозга неоднозначно и характеризуется множественностью физиологических эффектов. Поэтому в настоящее время построение модели, которая отражала бы зависимость между уровнем гормонов в организме и его влиянием на высшие функции мозга, представляется актуальным [35].
Исходя из вышеизложенного,целью работы стало исследование поведенческих особенностей крыс линий Wistar и WAG/Rij в зависимости от стадии эстрального цикла.
В соответствии с целью исследования, были были поставлены следующие задачи:
- Определить регулярность половых циклов у самок крыс линии Wistarи WAG/Rij.
- 2.Выявить уровни тревожности в динамике эстрального цикла у самок крыс линии Wistar и WAG/Rij.
- Исследовать уровень депрессивности в динамике эстрального цикла у самок крыс линии Wistarи WAG/Rij.
- Изучить нарушения когнитивных функций у самок крыс линии Wistarи WAG/Rijв динамике эстрального цикла.
Глава 1. Обзор литературы
1.1 Морфофункциональные изменения женской репродуктивной сֺиֺсֺтֺеֺмֺы
Яичники – это парный орган, который выполняет эндокринную (продукция половых гормонов) и репродуктивную (формирование женских половых клеток) функции. Они расположены асимметрично на заднем листке широкой связки в малом тазу и прикреплены нижним краем к короткой части мезовария. Укаждого яичника имеется две связки: собственная связкая ичника связывает яичник с маткой, которая заканчивается несколько ниже и позади маточной трубы и воронко-тазовая связка, которая направляется к боковой стенки таза от верхнего полюса яичника. В них и через мезоварий проходит основная часть нервов, лимфатических и кровеносных сосудов[44].
У яичника функционирование цикличное и, поэтому, его строение зависит от наличия беременности или от стадии эстрального цикла[26].
С наружней стороны, кроме области ворот, яичник покрыт эпителием, которого именуют по-разному покровной, поверхностный, зачатковый, или герминативный [13].
С поверхности яичник окружен плотной волокнистой соединительной тканью — белочной оболочкой, покрытой мезотелием. Остальная часть поверхности мезотелия обеспечена микроворсинками[4].
Яичник состоит из коркового и мозгового вещества. Корковое вещество расположено на периферии яичника и характеризуется наличием овариальных фолликулов. Мозговое вещество свободно от фолликулов и находится посередине яичника. В нем располагаются крупные органные сосуды, распадающиеся на более мелкие и, уходящие в корковое вещество,ветви. Специализированная рыхлая волокнистая соединительная ткань -стромальная часть этих веществ яичника и представляет собой производную мезенхимы[14].
Соединительнотканная строма является основой обоих веществ. В корковом веществе она представлена из межклеточного вещества и лежащих плотно фибробластов, а в мозговом веществе состоит из большого количества неупорядоченных эластических волокон, гладкомышечных клеток и огромного числа кровеносных сосудов. И преобладающая часть кровеносных сосудов сосредоточена в этом веществе [1,14].
Толщина коркового вещества постепенно возрастает от рождения до репродуктивного периода онтогенеза, а потом медленным темпом уменьшается. Толщина мозгового вещества –в период новорожденности самая наименьшая, а в период старости -наибольшая [50].
Корковое вещество состоит из веретеновидных тесно расположенных клеток, напоминающих набухшие фибробласты, и межклеточного вещества, которого очень мало. Но его тонкая внешняя зона представляет собой полосу коллагеновой стромы, которая содержит относительно мало клеток. Внутренняя зона коркового вещества яичников состоит из фолликулов, являющихся основной структурно-функциональной единицей [9].
Фолликулы подразделяются на примордиальные, первичные, вторичные и третичные(Граафовы), согласно Международной гистологической номенклатуры(1999). Их развитие происходит по схеме: примордиальный → первичный → вторичный → третичный [21]. Начало развития примордиальных фолликуловспиралевидных артерий и примордиальных фолликулов отмечается с периода полового созревания. В репродуктивный период фолликулы расположены в строме коркового вещества, примордиальные – напериферии, а зреющиев более глубоких зонах[25,51].
Примордиальный фолликул состоит из одного слоя плоских фолликулярных клеток ооцита первого порядка. Ооцит — ключевой элемент, основа функционального и структурного единства фолликулярного комплекса [49].
В примордиальных фолликулах гранулезные клетки небольших размеров. В дальнейшем увеличивается число и размеры гранулезы. Они становятся кубическими, цилиндрическими, формируют несколько слоев [27,28].
На начальных стадиях фолликулогенеза изменение размеров ооцита пропорциональноросту примордиального фолликула [33]. Первичные фолликулы формированы ооцитом первого порядка, который окружен немногочисленными слоями фолликулярных клеток в виде формы цилиндра или куба. Как только созревает фолликул, его окружают слоями веретенообразные клетки и увеличиваются в объеме, формируясь эпителиоидные. На периферии остаются слои веретенообразных клеток, которые потом сливаются со стромой.
Вторичный фолликул окружен наружней и внутренней текой, полость выстлана гранулезными клетками. В яичнике одновременно развиваются несколько полостных фолликулов, нодоминантным становится только один [12,13,24].
Он заполняется фолликулярной жидкостью по мере роста, оттесняет яйцеклетку к одному из полюсов, и превращается в граафов пузырек. В яйценосном холмике располагается яйцеклетка. С верхней стороны она покрывается лучистым венцом, блестящей и зернистой оболочками. [1,12,26].
Яйцеклетка с фолликулярной жидкостью выбрасывается в результате разрастания Граафова пузырька. На месте фолликула после овуляции и выброса яйцеклетки образуется желтое тело [12].
При оценке этапов развития фолликула по основным морфологическим признакам (форма и количество слоев фолликулярных клеток, наличие лакун) выделяют такие группы, как:
-вступившие в рост однослойные фолликулы,
-однослойные фолликулы с примордиальными ,
Рост и развитие фолликулов делятся на стадии:
1) образование пула растущих фолликулов;
2) рост до стадии антрального фолликула 4-го класса или базальный рост;
3) формирование и развитие доминантного фолликула.
Зависимость первых двух этапов от гипофизарных гонадотропинов является непрямой и опосредована факторами внутри яичников, а заключительный этап регулируется гипофизом напрямую [12]. Роль факторов клеточной адгезии, которые отвечают за межклеточные взаимодействия, несомненна. Интегрины α 3,6, иβ принимают участие в миграции первичной половой клетки [47]. Важна роль Е-кадерина и Р-кадерина в агрегации клеток при колонизации урогенитальных гребешков,экспрессирующиеся в предшественниках клеток Сертоли [46]. Факторы клеточной адгезии коннексинов играют важную роль в установлении взаимодействия ооцита и окружающих соматических клеток,а также в образовании фолликулов. По литературным данным, грызуны, у которых дефицит по коннексину СХ43, имеют серьезные нарушения в образовании примордиальных фолликулов [56]. Р-селектин играет важную роль на ранних этапах диапедеза лейкоцитов из кровотока во время воспаления [55].
Мозговое вещество яичников образовано рыхлой соединительной тканью. Вокруг нервных стволиков и сосудов нередко определяются маленькие пучки из полигональных или округлых клеток эпителиоидного вида. Эти клетки — рудиментарные остаткигонад, проходящих примитивную двуполую фазу развития. Они напоминают интерстициальные элементы яичек из-за выработки стероидов. Фолликулоциты гранулезной оболочки, текоциты внутренней теки, текоцитоподобные клетки стромы являются стероидпродуцирующими клетками яичников [44].
В литературных источниках нет единого мнения о строении стромы яичника. По данным Боровой Т. Г. (1993), строма яичников представляет собой самый «простой вариант»рыхлой соединительной ткани, практически единственными клеточными элементами, которой в корковом и мозговом веществе являются фибробласты и фиброциты. Помимо этого, в мозговом веществе присутствуют тучные и гладкомышечные клетки, небольшое количество лейкоцитов разной формы. По данным других исследователей строма она представлена двумя видами клеток:
1. веретенообразные, со скудной цитоплазмой, похожие на фибробласты, располагающиеся в плотной сети волокон с различными количествами коллагена;
2. полигональные, с эозинофильной цитоплазмой, свойственными стероидопродуцирующим клеткам ультраструктурными особенностями, богатые липидными включениями [8].
На основании современных представлений, гиалиноз артерий является основным морфологическим признаком стареющей гонады, при этом,вены компенсаторно расширяются. Инволюционный тип строения яичника характеризуется крупнобугристойповерхностьюоргана, склероз стромы, уплотнение белочной оболочки, малочисленность различных типов фолликул, наличие множества белых и фиброзных тел [28].
Овуляция –это динамическое явление, которое характеризуется воспалительным выбросом макрофагами цитокинов [56]. Однако, точной роли воспалительных и иммунологических событий в овуляции остается невыясненной.
Моноциты, макрофаги,ацидофильные гранулоциты, и нейтрофилы, инфильтрируют область, окружающую доминирующий фолликул перед овуляцией. Проникновение лейкоцитов, особенно нейтрофилов, наблюдается в theca слое яичника у человека[52] и крысы непосредственно перед овуляцией. Стромальные макрофаги могут быть вовлечены в регуляцию фолликулярного роста и разрыва, регуляцию эстрального цикла [59].
Только ограниченное число примордиальных фолликулов в яичниках млекопитающих растѐт и дифференцируется, достигая стадии доминантного фолликула и впоследствии овулирует. 99% фолликулов в яичнике подвергаются атрезии на различных стадиях развития. В репродуктивном возрасте атрезия совместно с формированием доминантного фолликула является одним из проявлений физиологии овариальной функции и фолликулогенеза: в ходе роста фолликулы соревнуются за лидерство, и поступательное развитие продолжают только те из них, которые способны обеспечить оптимальное морфофункциональное состояние собственного гистиона. В течение жизни в яичниках человека прогрессивно сокращается содержание половых клеток. Сравнительно небольшой процент этой убыли связан с созреванием и овуляцией ооцитов в репродуктивном периоде, и значительно больший — с атрезией овариальных фолликулов и гибелью половых клеток, происходящими практически в течение всей жизни. Апоптоз – фундаментальный механизм фолликулярной атрезии и постовуляторного регресса у млекопитающих[61].
Со временем наступления половозрелости половые органы самки подвергаются циклическим изменениям, протекающим в организме при участии нейроэндокринных стимулов и находящихся под влиянием факторов внешней и внутренней среды,которые, вероятно, оказывают влияние на формирование женских половых органов[28].
В результате большого количества антропогенных влияний число экологически зависимых патологических нарушений в женской репродуктивной системе растет. Как уже известно, по данным Губаревой Л. И. (2001), в яичниках крыс, которые подверглись воздействию сульфида и кадмия, значительная часть фолликулов на разных фазах созревания погибают. Следовательно,яичник- этоуникальный орган, где происходит постоянное изменение микроциркуляторного русла в течение жизни женщины, перестройка компонентов внеклеточного матрикса, транспорт лейкоцитов и тучных клеток, рости развитие различных функциональных структур: малых, средних, больших и атретических фолликулов, жѐлтых тел. В соответствии с данными литературы можно прийти к выводу, что генеративная и эндокринная функции яичников регулируются сложным комплексом гуморальных показателей[17].
1.2 Понятие эстрального цикла и его этапы
Под эстральным циклом понимается морфологические изменения, которые происходят циклически в генеративной системе самок, во время которого происходит созревание гамет и их выход в брюшную полость во время овуляции. Регуляцию эстрального цикла обеспечивают гормоны гипоталамо-аденогипофизарной конструкциии женской половой железы. Эстральный цикл делится на четыре фазы: проэструс (предтечка), эструс (течка), метаэструс (послетечка) и диэструс (межтечка). Каждая из фаз характеризуется определенным функциональным и клеточным составом слизистой влагалища, а также его гладко-мышечного слоя [8].
Подготовительной фазой эстрального цикла является проэструс. Происходит усиление продукции гонадотропоцитами аденогипофиза ФСГ под влиянием либеринов гипоталамуса. На данном этапе они переходят в фазу разрастания и начинается выработка эстрогенов. Их основной целью является матка. На данной фазе наблюдаются следующие изменения – слизистая оболочка начинает набухать и утолщаться, количество железистых клеток резко увеличивается, который начинают образовывать жидкий секрет, при этом происходит углубление желез и их разветвивание [36].
Следующая фаза – это фаза эструса. В крови по понятию обратной связи начинается нарастание концентрация эстрогенов,повышение продукции в гипоталамусе статинов. При этом происходит подавление секреции ФСГи лю-либеринов, следовательно,секреция лютеинизирующего гормона (ЛГ) начинается в гонадотропоцитах аденогипофиза . В результате того, что произошел рост концентрации в крови ЛГ, разрываются стенки зрелого фолликула в женской половой железе. Происходит овуляция, следствием чего в крови скапливаются эстрогены. Особенностью эструса является видовая специфика. В это время из наружных половых органов обильно выделяется слизь, когда матка готова принять зародыш [36].
Третьей фазой является метаэструс. Во время данной фазы в том месте, где лопнул зародыш образовывается желтое тело, основной целью которого является выработка прогестерона. Его основной целью является матка, в которой уже наблюдаются выраженные изменения в эндометрии. Эндометрий имеет наибольшую величину в периоде роста желтого тела. При этом железы становятся более глубокими и развлетленными. Гландулоциты начинают выделение густого секрета, который насыщен глюкогеном. В сосудах эндометрия наблюдается переполнения крови. Матка находится на стадии ожидания зародыша [36].
Последней стадией эстрального цикла является диэструс. В случае, если желтое тело не оплодотворилось, оно начинает увядать. Количество прогестерона в крови снижается, и описанные ранее процессы начинают протекать в обратном порядке.
Таким образом, в аденогипофизепродукции ЛГ и ФСГ протекают не в одно и то же время, в связи, с чем в яичнике секреция эстрогена последовательно сменяется продукцией прогестерона и, наоборот, в результате чего в половой системе самок протекают циклические изменения.
1.3 Эстральный цикл у крысы
Интервал эстрального цикла у мышей и крыс составляет 4-6 дней. Эстральный цикл продолжатся до наступления беременности. Во время беременности, протекающей 19-20 дней деятельность полового аппарата не наблюдается. Укрыс наступает течка на следующий день. После этого наступление эстрального цикла происходит только после завершения периода лактации [19].
Если процесс спаривания не закончился оплодотворением, либо в результате проведенного эксперимента наблюдается ложная беременность, то в матке и влагалище наблюдается изменения, длительность которых составляет 11 суток. значительные изменения влагалищных мазков и эпителия матки на морских свинках показали Stockard и Papanicolaou .
Когда влагалище находится в стадии покоя, оно покрыто слоями плоского эпителия. Вагинальные мазки содержат большой уровень лейкоцитов и малое количество эпителиальных клеток с ядрами.
В период течки клетки верхнего слоя слизистой оболочки влагалища отшелушиваются и происходит потеря ядер. В пробах мазка, которые были сделаны во время течки, находится достаточно большое количество данных чешуйчатых клеток. При этом наблюдается усиление секреции влагалищных желез.
Когда наступает стадия после течки видно, что лейкоцитарная инфильтрация стенки влагалища исчезает на стадии покоя. После чего в мазках можно наблюдать не большое количество клеток эпителиальных и большое число лейкоцитов.
Те изменения, которые были описаны, сопровождаются теми или иными видоизменениями матки.Во время предтечки она растет, железы ее расширяются и начинают выделять жидкость, растягивающая орган. после происходит дегенерация маточного эпителия и лейкоцитарная инфильтрация эндо- и миометрия. После этого лейкоциты исчезают, эпителий восстанавливается и у эндометрия образуется способность формирования децидуальных клеток при последующем спаривании.
1.4 Поведенческие эффекты женских половых гормонов.
Женский половой гормон — эстроген, вызывающий у женского пола ощущение благополучия и удовлетворения, обуславливающий манеру поведения (стремление нянчить свое потомство и защищать свое гнездышко). Эффект эстрогенов на ЦНС делится геномный и негеномный. При первом влияние действия женских половых гормонов проходит через синаптические контакты и мозговые структуры с помощью специфических рецепторов. Стероиды, которые циркулируют в крови, влияют на функции комплекса, участвующего в организации висцеральных, мотивационных и эмоциональных реакций организма. Геномные эффекты постоянны и контролируют нейронную архитектонику. Негеномные эффекты эстрогенов затрагивают механизмы нейропередачи: изменение концентрации и доступности серотонина и норадреналина, увеличение времени деградации моноаминоксидазы (фермент, метаболизирующийи усиливающий транспорт серотонина), вытеснение триптофана из связанного состояния с альбумином плазмы, что повышает число свободного триптофана, поступающего в мозг, где и превращается в серотонин. Подтверждение важности этих моноаминных нейротрансмиттерных систем в регуляции настроения есть заслуга влияния женских половых гормонов на обмен норадреналина и серотонина в центральной нервной системе. Они оказывают серотонинэргический эффект (повышение активности серотонина). Ряд научных исследований указывают на положительный эффект применения заместительной гормональной терапии на настроение и чувство общего благополучия у женщин в постменопаузе, включая женщин с отсутствием психологических симптомов. Также выявлено, что женские половые гормоны купируют такие симптомы, как тревогу, снижение самооценки и депрессивное настроение. Эстрогены оказывают влияние на ЦНС, и на психическую сферу в частности, посредством нескольких механизмов. Одним из них является прямое психотропное (психостимулирующее и антидепрессивное) действие эстрогенов за счет их серотонинэргического и адренергического эффектов. С другой стороны, вследствие уменьшения вазомоторных симптомов (приливов жара, потливости) улучшается общее самочувствие и психическое состояние пациенток («домино-эффект») [16].
Существуют еще гормон, влияющий на состояние женщины,-прогестерон.Известно, что среди хорошо изученных эндокринных и репродуктивных эффектов, он играет главную роль в регуляции пластических процессов в головном и спинном мозге, периферической нервной системе.Также установлено, что он синтезируется в одинаковых концентрациях у мужского и женского пола в глиальных клетках и нейронах центральной и периферической нервной системы. Его метаболиты связываются с теми же рецепторами, которые опосредуют эффекты противотревожных средств (бензодиазепиновых транквивлизаторов) – ГАМК-А/бензодиазепиновым рецепторным комплексом, и оказывают психотропное действие на организм женщины, что нашло свое место в лечении некоторых форм депрессии, агрессии, мигрени. Активация ГАМК-рецепторов ведет также к улучшению питания нервной ткани. Когда в конце месячного цикла резко снижается секреция прогестерона, ослабляется трофическая функция ЦНС, что проявляется в повышенной утомляемости, раздражительности, расстройствами настроения, среди которых преобладают чувство беспричинной тревоги. С резким падением продукции прогестерона по окончанию беременности связано и повышение частоты депрессивных состояний в послеродовом периоде. Производные прогестерона в гормональных контрацептивах оказывают гипнотический эффект, который используется для лечения предменструального синдрома и психологического стресса [32].
Специфической общепризнанной поведенческой реакцией на стресс является груминг .По данному параметру можно оценить нейрогуморальные изменения в организме. Анализируя характер влияния женских половых гормонов различного происхождения на груминговую активность крыс с исходно разным уровнем активности, выявлено, что естественный женский стероид эстрон стимулирует груминг у низкоактивных крыс и подавляет у средне- и высокоактивных животных. Синэстрол, напротив, не повлиял на груминговое поведение стрессированных крыс. Данный факт указывает на анксиогенное влияние эстрона на низкоактивных в контроле крыс и анксиолитическое – на средне- и высокоактивных стрессированных самцов белых крыс[37].
Учитывая полученные данные, было сделано предположение, что изменения в эстральном цикле и половом поведении обусловлены с дисбалансом эстрогенов и гестогенов в крови связанным с дефицитом магния. В этой связи сочли целесообразным изучить и процессы зачатия у этих самок [29].
1.5 Поведенческие особенности крыс линии WAG/Rij
В настоящее время в практике поведения клинических исследований используются животные различных линий. Крысы нужны для установления токсичности лекарственных веществ и ядов, широко используются при изучении вопросов питания, проведения биологической стандартизации гормональных препаратов для постановки научных исследований по витаминологии, физиологии, фармакологии, эндокринологии, биохимии и для изучения процессов поведения и других вопросов психологии. Но следует знать поведенческие реакции крыс разных линий.
WAG/Rij – инбредная линия крыс, выведенная из крыс Wistar Albino Glaxo в Великобритании (Wistar Albino Glaxo Laboratory, UK) в 1924г. Позднее эта линия крыс была ввезена в Нидерланды (REPGO Institute of TNO at Rijswijk, The Netherlands). WAG/Rij означает Wistar Albino Glaxo/Rijswijk (Саркисова К.Ю., 2005). Крысы линии WAG/Rij – результат скрещиваний более 130 поколений (Depaulis A., van Luijtelaar G., 2006).
Известно, что один из патогенетических механизмов эпилептической активности у крыс линии WAG/Rij является гипофункция дофаминергической системы мозга [50].
Несмотря на это, механизмы формирующегося снижения активности этой системы нераскрыты. Определено, что дефицит дофаминергической системы у крыс линии WAG/Rij может быть предопределен динамикой функционирования дофаминовых рецепторов [50,53].
Рецепторы D2- самый распространённый тип дофаминовых рецепторов. Они расположены в большем количестве в лимбической системе, в коре головного мозга, в среднем мозге. Стратегия приспособительного поведения зависит от уровня и функционирования данных рецепторов, которая проявляется в двух формах — активная и пассивная [40].
Таким образом, исследование тревожно-депрессивных состояний в зависимости от уровня половых стероидов при изучении эпилепсии и других патологий нервной системы является актуальным и имеет большое практическое значение. Однако эта область исследований мало изученная и требует детальное выяснение взаимосвязи поведения, эмоциональной реактивности и тревожности с уровнем стероидных гормонов в различных отделах мозга для понимания их нейроактивных и нейроэндокринных функций.
Глава 2. Материал и методы исследования
2.1 Объекты исследования
Эксперименты проводились на крысах линии Wistar (n=20) и WАG/Rij (n=20), которые содержались в условиях вивария кафедры физиологии человека и зоологии Башкирского государственного университета, характеризующихся постоянством комнатной температуры (20-22 °C) и уровнем влажности. Животные находились в стандартных пластмассовых клетках по 3-4 особи. Условия содержания для всех крыс были одинаковыми. Клетки, кормушки, поилки и весь необходимый инвентарь систематически обрабатывались с использованием специальных дезинфицирующих средств. В качестве подстилки использовали опилки и мелкую стружку, которые менялись по мере загрязнения, но не реже 1 раза в неделю.Питались в основном зерновой смесью, свежими овощами и творогом.При работе с крысами полностью соблюдались международные принципы Хельсинкской декларации о гуманном отношении к животным (2000 год).
2.2 Методы исследования
2.2.1 Определение стадий эстрального цикла самок крыс методом влагалищных мазков
Мазки из влагалища крыс можно брали двумя способами: с помощью пипетки или ватной палочки. Заранее следует заготовить достаточное количество палочек, так как каждую можно использовать только один раз. На рабочем столе должны находиться емкость с водой, нужное количество обезжиренных предметных стекол и восковой карандаш (маркер использовать не рекомендуется, так как при фиксации препарата надпись растворится). Обезжиривание обычно проводят длительным выдерживанием предметных стекол в растворе этанола с диметиловым эфиром в пропорции 1:1.
Перед взятием мазка ватная палочка погружается в чашку с водой, и капля воды переносится на предметное стекло. Прикасаться пальцами к тампону нежелательно, так как клетки эпидермиса кожи пальцев, попав мазок, могут нарушить микроскопическую картину. После этого намоченный конец ватной палочки осторожно вводят во влагалище и слегка там поворачивают. Вынув его из влагалища, размазывают захваченную ватой пробу в капле воды, нанесенной на предметное стекло. На стекле подписывают дату взятия мазка и номер крысы.
При систематическом взятии мазков у одних и тех же животных, предпочитают брать пробы глазной пипеткой, во избежание механического повреждения влагалища. Для этого набирается несколько капель воды в пипетку и, неглубоко вводя кончик во влагалище, 1-2 раза выжимают воду из пипетки и насасывают ее обратно. Взятая проба переносится на обезжиренное предметное стекло.
В условиях длительного эксперимента забор влагалищных мазков необходимо проводить в фиксированное время суток [30].
Неокрашенные пробы лучше рассматривать под микроскопом немедленно, так как по мере подсыхания изучение мазка становится более затруднительным. Считается, что более точно мазок распознается в фиксированном и окрашенном препарате [22].
С целью определения ритмичности функционирования яичников, ежедневно анализировали влагалищные мазки. По следующей методике проводили приготовление мазков и анализировали их. Влагалищные смывы брали тонкой глазной пипеткой, помещали на предметное стекло и рассматривали под микроскопом при увеличении в 300 раз в слегка затемненном поле [7]. Эстральный цикл крысы состоит из 4-х фаз: проэструс (предтечка), эструс (течка), метаэструс (послетечка) и диэструс (межтечка, или стадия покоя). Каждой стадии эстрального цикла соответствует определенный клеточный состав влагалищного мазка (табл.1).
После определения фазы полового цикла каждого животного результаты были подвергнуты обработке и анализу.
2.2.2 Тест «Открытое поле»
Самые разные поведенческие тесты используются для изучения количественных и качественных индексов поведения[22]. Одним из самых распространенных тестов, который используется в основном в составе комплекса тестов, является тест «открытое поле». Круглое открытое поле представляет собой арену 1,5 м в диаметре с высотой стенок 0,8 м, дно которой было расчерчено на сектора. В открытом поле выделяли 3 зоны, по которым далее производили оценку всех количественных показателей: центральная зона, зона середины (6 сегментов), периферическая зона (12 сегментов), (рис. 1).
 Рис. 1. Круглое открытое поле
Рис. 1. Круглое открытое поле
Таблица 1. Клеточный состав влагалищного мазка и средняя продолжительность стадий эстрального цикла.
| Стадия | Средняя продолжительность в часах | Состав содержимого влагалища |
| Проэструс | 12 ч | Слизь и лейкоциты исчезают,мазок почти исключительно состоит из эпителиальных клеток, слущенных со стенки поодиночно или небольшими группами. Все эпителиальные клетки примерно одинаковой величины,имеют овальную форму на окрашенном мазке видно, что все эпителиальные клетки содержат ядра. |
| Эструс | 27 ч | Одинаковые ороговевшие чешуйки — безъядерные ,ороговевшие клетки. Лейкоциты и эпителиальные клетки с ядрами совершенно отсутствуют. На этой стадии полового цикла нормально происходит спаривание. К концу стадии чешуйки образуют хорошо видимые беловатые скопления. |
| Метаэструс | 6 ч | Среди массы ороговевших клеток (чешуек) появляются лейкоциты. На короткое время могут появиться и эпителиальные клетки, так что в мазке встречаются все 3 типа клеток (чешуйки, эпителиальные клетки и лейкоциты). |
| Диэструс | Значительно варьирует | Значительное количество слизи. Основную массу клеток составляют лейкоциты |
Освещение производилось 2 лампами, мощностью по 60 Вт, которые располагались на высоте 1,5 м от дна камеры над центральными сегментами поля. Все процедуры связанные с тестированием в открытом поле проводились с 14.00 до 18.00.
Крыс помещали в центр открытого поля хвостом к экспериментатору и оценивали двигательную и исследовательскую активность животного. Двигательную активность определяли путем подсчета числа пересеченных животным сегментов. Сегмент считался пересеченным, если крыса переступала через линию передними и задними лапами. Общая двигательная активность вычислялась путем суммирования числа пересеченных квадратов в центре, в середине поля и по его периферии. Неподвижностью (сек.) являлся период замирания животного или пассивного бодрствования. Она характеризовалась полным отсутствием локомоции.При обработке эксперимента выделяли неподвижность до первого двигательного акта в поле – латентный период и неподвижность, зафиксированную в течение сеанса тестирования.
Исследовательскую деятельность определяли путем подсчета совершенных вертикальных стоек в различных зонах поля. За стойку принимали положение животного, когда оно отрывало передние лапы от пола, и оставалось лишь на задних лапах. При этом животное вытягивало шею, часто поворачивая голову, и принюхивалось. Общее количество совершенных стоек подсчитывалось при суммировании стоек, совершенных в центре, в середине и на периферии поля.
За эпизод груминга принимали все периоды времени, когда животное проявляло соответствующее поведение по уходу за собственным телом (умывание, вылизывание, вычесывание). Время, посвященное процессу чистки, фиксировалось по зонам, и затем вычислялся общий показатель длительности груминга. Общее число эпизодов груминга характеризовали его интенсивность. Под общим числом мы понимали количество совершенных актов чистки в различных зонах поля. Вегетативные показатели, определяющие эмоциональный статус животного, представляли из себя акты уринации и дефекации. Количество дефекаций и уринаций высчитывали по окончанию тестирования каждого животного. Данные за пятнадцать минут суммировали и заносили в таблицу,после вычисляли среднее значение за 5 дней тестирования. Стандартный режим регистрации соблюдался нами в течение всех 5 дней тестирования. Эксперимент проводился в одно и тоже время дня, в затемненной комнате с освещением центральной площадки поля, без постороннего шума и других раздражителей. После тестирования каждого животного поле тщательно обрабатывалось спиртовым раствором и промывалось водой.
2.2.3 Тест «Приподнятый крестообразный лабиринт»
«Приподнятый крестообразный лабиринт» (ПКЛ) представляет собой площадку, имеющую 2 закрытых (со стенками) и 2 открытых (без стенок) рукава, шириной 10 см и длиной 45 см каждый. Лабиринт располагался на высоте 80 см от пола. В центре расположенных крест-накрест рукавов лабиринта находилась открытая площадка ,размером 10*10 см [12].
 Рис 2. Установка «Приподнятый крестообразный лабиринт»
Рис 2. Установка «Приподнятый крестообразный лабиринт»
Животное помещали на данную площадку и в течение 5 минут регистрировали следующие параметры: число заходов на светлое и темное поле, время нахождения в каждом поле, количество совершенных стоек, неподвижность, частоту уринаций и дефекаций, количество свешивания в светлом рукаве , длительность груминга (сек.),количество эпизодов груминга.
Стандартный режим тестирования сохранялся нами для всех животных. Эксперимент проводился в одно и тоже время дня, без постороннего шума и других раздражителей. После тестирования каждого животного установка тщательно обрабатывалась спиртовым раствором и промывалась водой.
2.2.4 Тест «Вынужденное плавание»
 Рис 3. Установка теста «Вынужденное плавание»
Рис 3. Установка теста «Вынужденное плавание»
Источник