
Основные части тела муравья.
Основные части тела муравья.
Основными частями тела муравья являются голова, грудь (мезосома), стебелёк и брюшко. Стебелёк может состоять из одного (у большинства подсемейств) или двух члеников (у подсемейства Myrmicinae).
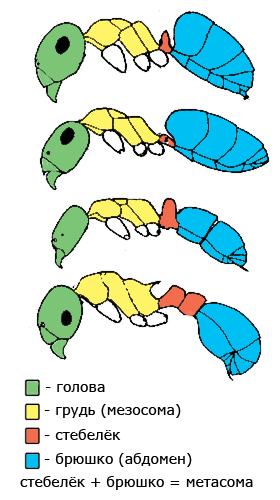
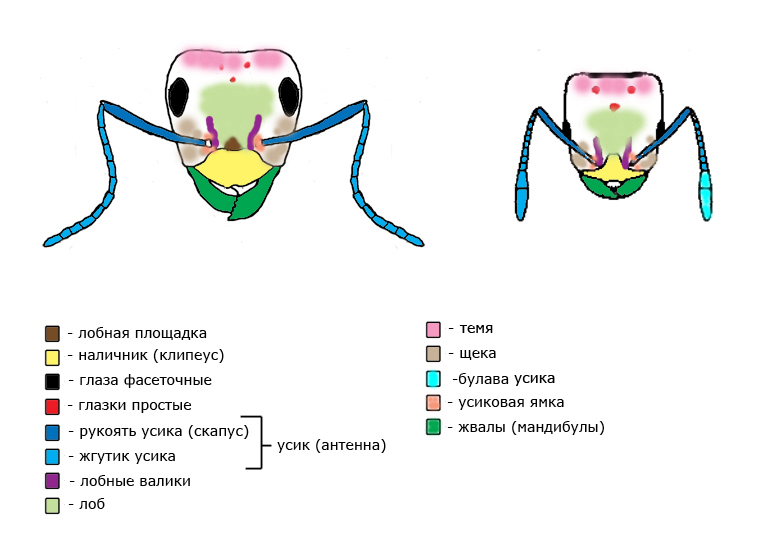
Голова состоит из головной капсулы, несущей глаза и простые глазки (если они есть), усики и ротовые части. Причленяется к груди. Усик состоит из рукояти (скапуса) и жгутика. Ротовой аппарат устроен таким образом, что муравей может поглащать только жидкую пищу. Твёрдую пищу могут потреблять личинки муравьев.
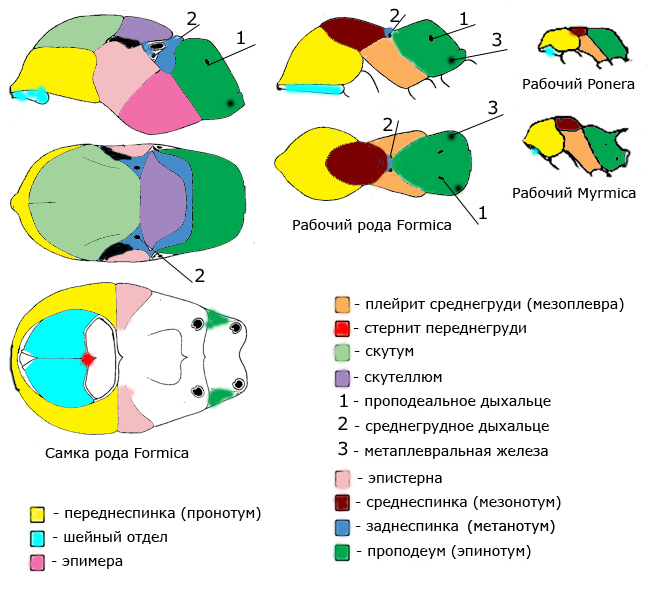
Грудь (мезосома) состоит из переднегруди, среднегруди и проподеума (эпинотума). грудь самок и самцов разделена более сильно. К их груди крепятся крылья. Грудь рабочих муравьев расчленена значительно меньше. Многие её части слиты, швы между ними исчезли. У некоторых видов на проподеуме (эпинотуме) есть шипы.
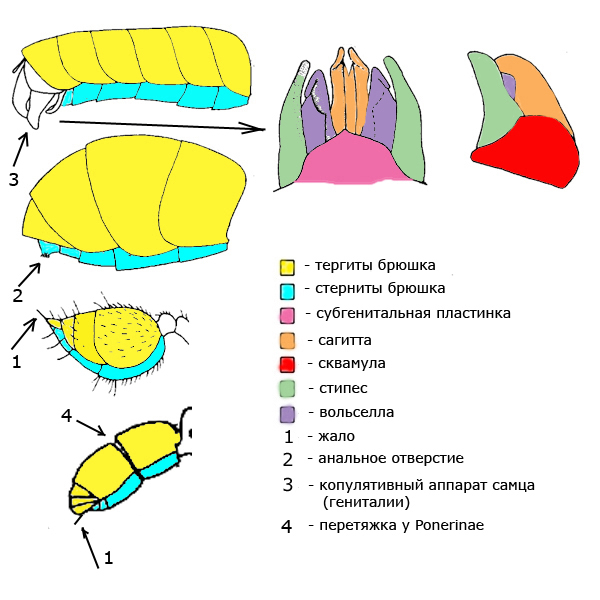
У муравьев имеющих стебелёк из одного членика брюшко состоит из 5 сегментов у самок и рабочих особей и 6 сегментов у самцов. У муравьев имеющих стебелёк из двух члеников соответственно 4 и 5 сегментов. Стебелёк и брюшко вместе образуют метасому. Кольцо сегмента состоит из двух полуколец: тергита сверху и стернита с нижней стороны. На конце брюшка самцов находится копулятивный аппарат (гениталии). У некоторых видов у рабочих особей и самок есть жало.
Передние ноги отличаются тем, что на них имеется специальное приспособление для чистки усиков и лапок с большой гребенчатой шпорой.
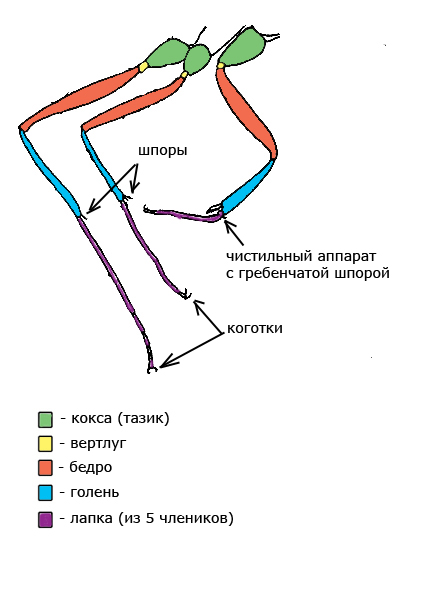
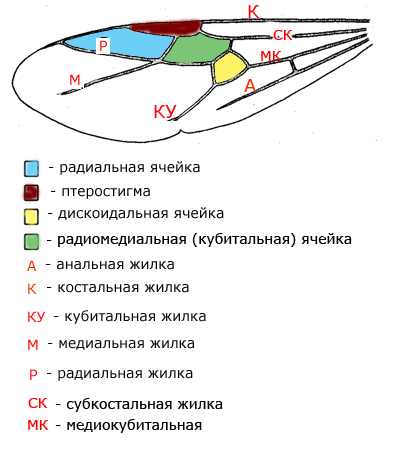
Крылья бывают только у самок и самцов. Крыльев всего четыре — два передних и два задних. После брачного лёта самка крылья сбрасывает. Схема переднего крыла показана ниже:
Вы можете почитать список названий частей тела, используемых в русскоязычной литературе (список рус.) и в англоязычной литературе (список англ.)
На данной странице использованы рисунки из работ:
Источник
Список названий частей тела муравья встречающихся в русскоязычной литературе.
Список названий частей тела муравья.
Абдомен – см. брюшко.
Антенна – см. усик.
Аролий – имеется у некоторых муравьев на кончиках лапок между коготков.
Ацидопора — выход ядовитой железы на кончике брюшка вытянутый обычно в трубочку окруженную венчиком волосков.
Бедро – часть ноги.
Брюшко (абдомен) – бывает округлым, вытянутым, сердцевидным. Верхние пластины брюшка называются тергитами, нижние – стернитами. Тергиты имеют отверстия дыхалец.
Вертлуг – небольшой членик ноги между коксой (тазиком) и бедром.
Верхняя губа – часть ротового аппарата муравья.
Вольселла — часть наружных половых органов самца.
Галеа – часть ротового аппарата муравья.
Гениталии – наружные половые органы самца.
Глаза – сложные фасеточные глаза муравья. Их два. Обычно состоят из множества фасеток. У некоторых видов могут состоять всего из нескольких фасеток. Есть виды муравьев вообще лишённые глаз.
Глазки – простые глазки из одной фасетки. Их три. У некоторых видов их имеют только половые особи. У некоторых видов их нет.
Голень – часть ноги.
Голова – состоит из головной капсулы, несущей глаза и простые глазки (если они есть), усики и ротовые части. Причленяется к груди.
Грудь (мезосома) – центральная часть тела муравья.
Гуларная пластина – пластина на нижней стороне головы между жвалами. В американской литературе гулой часто называют всю нижнюю поверхность головы.
Жвалы (мандибулы) — верхние челюсти, чаще массивные, в виде широких пластинок, с большим количеством зубчиков на жевательном крае, с выпуклым внешним и вогнутым внутренним краем, реже саблевидные без зубцов.
Заднеспинка (метанотум) – часть груди. Хорошо различима у самок и самцов. У рабочих особей слита с эпинотумом.
Затылок – область головы от темени до затылочного отверстия.
Затылочный край головы – задний край головы, если смотреть на неё точно сверху.
Кардо – часть ротового аппарата муравья.
Клипеус – см. наличник.
Кокса (тазик) – верхняя часть ноги.
Копулятивный аппарат – см. гениталии
Лапка – нижняя часть ноги. Состоит из пяти члеников. Первый членик самый длинный. На конце лапки два коготка.
Лациния – часть ротового аппарата муравья.
Лоб – часть головы без четкой границы. Находится между лобными валиками, граничит со щёками по бокам и теменем сверху.
Лобная бороздка — бороздка, отходящая вверх от лобной площадки. Бывает длинной (до переднего глазка), короткой или отсутствует.
Лобные валики — бороздки, которые бывают прямыми или изогнутыми, параллельными или расходящимися в стороны, длинными или короткими. У некоторых видов лобные валики очень короткие или совсем не выражены.
Лобная площадка (лобный треугольник) – треугольная пластина между усиками над наличником.
Максиллы – часть ротового аппарата муравья.
Мандибулы – см. жвалы.
Мезоплевра – бока среднегруди у рабочих особей.
Мезонотум – см. среднеспинка.
Мезосома – см. грудь.
Метаплевральная железа – парные железы, выделяют антибиотические вещества, а иногда также феромоны тревоги и репелленты для защиты от врагов.
Метасома – стебелёк и брюшко взятые вместе.
Метанотум – см. заднеспинка.
Наличник (клипеус) – представляет собой квадратную, треугольную, полукруглую или трапециевидную пластинку. Передний край наличника прямой, слабовыпуклый или вогнутый, иногда с выемкой на середине или с зубчиками по бокам; задний край угловатый. Поверхность наличника чаще выпуклая, реже с продольным килем, иногда с несколькими продольными валиками или поперечным вдавлением. По бокам наличника в верхней части расположены наличниковые ямки.
Нижняя губа – часть ротового аппарата муравья. Состоит из лорума, прементума с парой нижнегубных щупиков, параглосс и язычка, образованного слившимися глоссами.
Нижнечелюстные щупики – часть ротового аппарата муравья.
Переднеспинка (пронотум) – передняя часть груди муравья.
Плейрит (мезоплевра) – часть груди.
Пронотум – см. переднеспинка.
Проподеум (эпинотум) – задняя часть груди.
Проподеальное дыхальце – парные дыхальца на проподеуме (эпинотуме).
Псаммофор – специальное приспособление для переноски сухой почвы, распространённое у пустынных муравьев. Образован длинными отстоящими волосками (аммохетами), расположенными на нижней поверхности головы.
Птеростигма – утолщение у передней кромки крыла.
Сагитта — часть наружных половых органов самца.
Сенсиллы – отстоящие волоски на последних члениках усика. Являются органами осязания.
Сквамула — часть наружных половых органов самца.
Скутеллюм – часть груди самок и самцов
Скутум – часть груди самок и самцов
Среднегрудное дыхальце – парные дыхальца, расположенные на мезопроподеальном шве.
Среднеспинка (мезонотум) – часть груди.
Стебелёк – состоит из одного членика (петиолюса) или двух члеников (петиолюса и постпетиолюса). Если состоит из одного членика, то часто развивается в поперечную лопасть, носящую название чешуйки. Чешуйка может быть вертикальной или наклоненной вперед. Реже петиолюс может быть цилиндрическим (Tapinoma) или узловидным (некоторые Cataglyphis). Напротив, у видов с 2-члениковым стебельком форма петиолюса бывает, чаще всего, узловидной, с развитым узелком различной формы и, как правило, удлиненной передней цилиндрической частью, реже цилиндрической (Myrmecina). Постпетиолюс у этих видов имеет более или менее шаровидную форму. Стернит переднегруди – часть груди.
Стернит среднегруди – часть груди.
Стипес – часть ротового или полового (копулятивного) аппарата муравья.
Субгенитальная пластина – часть наружных половых органов самца.
Тазик – см. кокса.
Темя – поверхность головы сразу за глазками. За теменем до затылочного отверстия находится затылок.
Усик (антенна) – у муравьев коленчатый. Усик состоит из рукояти (скапуса) и жгутика. Последние членики жгутика у некоторых видов увеличены и образуют булаву.
Усиковые ямки – углубления у края наличника. Место причленения усиков.
Шейный отдел – образуется за счет слияния шейных склеритов и плейритов переднегруди. Подвижно сочленяется с переднеспинкой.
Шпора – отросток на голени ноги.
Щека – участок головы от нижнего края глаз до места прикрепления жвал (мандибул).
Эпимера – боковые части груди самок и самцов.
Эпинотум – см. проподеум.
Эпистерна – боковые части груди самок и самцов.
Источник
Воспользуйся поиском! Ты найдешь ответы на 99% своих вопросов.
Основное меню
Навигация
Сервисы
Сейчас на сайте
Просто реклама
Анатомическое строение муравья

| 1 — Нижняя губа; 2 — Предротовая камера; 3 — Глотка; 4 — Пищевод; 5 — Зоб; 6 — Провентрикулюс; 7 — Средняя кишка; 8 — Пилорический отдел средней кишки; 9 — Тонкая кишка; 10 — Прямая кишка; 11 — Анальное отверстие; 12 — Максиллярная железа; 13 — Нижнечелюстные железы; 14 — Глоточная железа; 15 — Слюнная железа. |
Органы пищеварения у муравьев разделяются на предротовую камеру и собственно пищеварительный тракт.
Предротовая камера – это сферическая полость, расположенная над нижней губой и под глоткой. Она служит приемником для жидкой и полужидкой пищи, а также для различных остатков после чистки тела. В предротовой камере происходит «сортировка» пищи – всё съедобное попадает в рот, а несъедобные частицы выделяются в виде комочков, имеющих форму камеры.
Пищеварительный тракт состоит из переднего, среднего и заднего отделов. Передний отдел у взрослых муравьев состоит, пищевода, зоба и провенрикулюса. За несколько вздутой глоткой, находящейся в передней части головы и открывающейся ротовым отверстием, следует длинный пищевод, проходящий через всю грудь насекомого. Зоб, слепой отросток пищевода, у многих муравьев может сильно раздуваться. По образному выражению, зоб – это «общественный желудок» муравьев. Пища, хранящаяся в нем, распределяется среди всего населения гнезда.
Интересно строение провентрикулюса, или жевательного желудка, — последнего отдела передней кишки. Как показали исследования Эйзнера и его соавторов (Eisner 1957; Eisner, Brown, 1958; Eisner, Happ, 1958), сложное строение этого отдела имеет большое функциональное значение. У примитивных муравьев(например муравьи подсемейства Myrmicinae) прохождению пищи из зоба в желудок препятствует только кольцевой мышечный сфинктор, поэтому зоб не может в полной мере функционировать как «общественный желудок». Трофаллаксис у этих муравьев развит слабо, длительное время хранить пищу в зобу они также не способны. У Formicinae и Dolichoderinae появляются специальные приспособления, позволяющие без мышечных усилий удерживать пищу в зобу. Провентрикулюс у этих муравьев жесткий и склеротизированный, а чашеобразные и куполообразные структуры образуют клапаны, которые автоматически не пропускают пищу в желудок.
Во вздутой средней кишке, которую обычно называют желудком, происходит основное переваривание пищи. По данным Эйра(Ayre, 1963) у Camponotus herculeanus только здесь выделяется протеаза и преимущественно здесь же – липаза. Из ферментов, разлагающих углеводы, он обнаружил лишь небольшое количество инвертазы. Однако у Formica polyctena в этом отделе активно разлагаются мальтоза, сахароза и мелитоза и слабее мелобиоза, раффиноза, трегалоза и крахмал(Graf, 1964).
Возле средней кишки Serviformica локализуются симбиоциты – сильно видоизмененные клетки, содержащие симбиотические бактерии.
Задняя кишка подразделяется на три отдела: пилорический отдел, тонкую кишку и прямую кишку, или ректум. Последняя сильно вздута, снабжена мошной мускулатурой и открывается в анальную трубочку.
У Formica имеются следующие парные железы, принимающие участие в процессе пищеварения: максиллярные (нижнечелюстные), слюнные (лабиальные) и глоточные. У F. polyctena экскреты этих желез разлагают следующие углеводы: мальтозу, сахарозу, мелитозу, мелобиозу, раффинозу, трегалову и крахмал (Graf, 1964).
Максиллярные железы открываются в глотку. У С. herculeanus они выделяют главным образом инвертазу и в меньшей степени — амилазу, т.е. ферменты, переваривающие углеводы (Ayre, 1963).
Лабиальные (слюнные) железы располагаются в груди и гомологичны прядильным железам личинок. Их две, но протоки этих желез сливаются вместе и образуют один непарный проток, открывающийся в нижней губе. У F. rufa на каждом из парных протоков перед слиянием имеются слепые отростки, способные раздуваться, служащие для хранения экскрета (Meinert, пo Wheeler, 1910). У С. herculeanus основной фермент, выделяемый этой железой — амилаза (Ауге, 1963).
Как показали исследования Гессвальда и Клофта (Gosswald, Kloft, 1957—1960) с применением радиоактивного фосфора, экскрет лабиальных желез служит для кормления цариц и личинок половых особей. Меченый фосфор из желудка через 24 часа попадает в эти железы, а затем уже экскрет распределяется в гнезде.
Глоточные (фарингеальные или, правильнее, постфарингеальные) железы муравьев не гомологичны глоточным железам других перепончатокрылых, например, пчел (Otto, 1958b). У С. herculeanus экскрет глоточных желез содержит небольшое количество липазы и следы амилазы (Ауге, 1963). Опыты с радиоактивным фосфором показали, что у Formica из железы этот экскрет поступает в зоб и затем распределяется между всеми особями гнезда (Naarman, 1963).
Кроме перечисленных желез, с ротовым аппаратом муравьев связаны парные мандибулярные (челюстные) железы, открывающиеся в основании жвал. К процессу пищеварения эти железы, по-видимому, не имеют отношения. Считается, что они выделяют вещества, используемые для склеивания частиц почвы при постройке гнезда или для изготовления картона (Donisthorpe, 1915). У ряда видов из подсемейств Myrmicinae и Dorylinae эти железы выделяют пахучие вещества — торибоны (Wilson, 1963b).

M — мальпигиевы сосуды
Органы выделения представлены у муравьев малышгиевыми сосудами, впадающими в пилорический отдел задней кишки. Функцией их является выведение из организма конечных продуктов обмена веществ, главным образом мочевой кислоты.
Хитиновый наружный скелет служит основой, к которой прикрепляются поперечнополосатые скелетные мышцы. У рабочих строение мышечной системы проще, поскольку у них отсутствует летательная мускулатура, имеющаяся у самцов и самок. Однако у последних она, после сбрасывания крыльев, резорбируется и идет на образование экскрета, которым выкармливаются личинки .
Дыхательная система муравьев, как и у подавляющего большинства других насекомых, трахейная. Трахеи открываются наружу дыхальцами, или стигмами . Дыхальца имеются между среднегрудью и эпинотумом (заднегрудные), на эпинотуме, на стебельке у основания чешуйки и на каждом из сегментов брюшка.
Гемолимфа («кровь») муравьев — бесцветная жидкость. Она циркулирует по телу насекомого благодаря работе спинного сосуда («сердца») — мускулистой трубки, проходящей вдоль всей спинной поверхности тела.
Центральная нервная система

1a — надглоточный ганглий; 1b — подглоточный ганглий; 2 — грудные нервные узлы; 3 — брюшная нервная цепочка.
Центральная нервная система насекомых состоит из ряда ганглиев, связанных между собой. У Formica имеются следующие ганглии: надглоточный, подглоточный, три грудных (соответствующие каждому сегменту груди) и несколько небольших брюшных.

Сравнительные размеры и форма надглоточного ганглия
рабочего(1), матки(2) и самца(3) Serviformica fusca.
Наиболее важной частью является надглоточный ганглий, или «мозг» муравьев, в котором образуются временные связи. Объем «мозга» относительно наибольший у рабочих, меньше у самок и самый маленький у самцов . По данным Маршала (Marchal, по Шовену, 1953), объем мозга Formica составляет 1/280 объема тела, у Dytiscus это отношение равно 1/4200, у Ichneumon — 1/400 и у медоносной пчелы – 1/174.
Временные связи образуются у муравьев в грибовидных телах, являющихся аналогом коры головного мозга позвоночных. Размеры грибовидных тел муравьев связаны со способностью различных видов к образованию условных рефлексов (Brun, 1959). У рабочих Formica (Marchal, по Шовену, 1953) грибовидные тела составляют 1/2 объема мозга, у самок они относительно меньше, а у самцов совсем маленькие. Для сравнения отметим, что у медоносной пчелы, несмотря на то, что мозг их относительно больше, грибовидные тела составляют всего 1/15 размеров мозга.

Органы зрения представлены большими фасеточными глазами и тремя простыми глазками,
имеющимися у всех каст. Функция глазков пока не очень ясна. Имеются данные (Homann, 1924),
что Formica с глазками, покрытыми непрозрачным лаком, ведут себя как слепые. Сложный глаз
состоит из большого количества отдельных омматидиев. Для разрешающей способности глаза
большое значение имеет угол зрения отдельных омматидиев. Например, у пчелы этот угол около 1°, а у уховертки — 8°, так что там, где уховертка видит только одну точку, пчела различает 64 (Шовен, 1953). У F. rufa угол зрения отдельного омматидия рабочего 3,5 , но насекомое может различать сферу при телесном угле 2,5° (Homann, 1924). Еще старыми наблюдениями Леббока и Фореля (Леббок, 1898; Forel, 1886а) было установлено, что муравьи собирают своих личинок на границе видимого и инфракрасного света (800 ммк), но избегают темной для нас зоны ультрафиолетовых лучей (380—330 ммк). Они уносят личинок под сосуд с сероуглеродом, поглощающим ультрафиолетовые лучи, но прозрачным для нас, предпочитая его экрану, зачерненному окисью никеля, пропускающей ультрафиолетовые лучи, но непрозрачной для видимых. Наиболее активно стимулирует у муравьев перенос личинок зона от 600 до 575 ммк (желтый свет) (Эббот, по Шовену, 1953). В последние годы (Vowles, 1950) доказано, что муравьи, так же как и пчелы, способны воспринимать направление колебаний поляризованного света.

Запах муравьи воспринимают жгутиком усиков. Муравьи великолепно различают тончайшие оттенки запаха, непостижимые для нас. Однако мнение старых авторов (Леббок, 1898; Forel, 1921 и др.), что муравьи способны различать по запаху даже направление следа, было опровергнуто опытами Шовена (Шовен, 1960).
Органы вкуса муравьев расположены на жгутиках усиков, на нижней губе и, по-видимому, на максиллах. На жгутиках усиков, возможно, органами вкуса являются многочисленные здесь пластинки, пронизанные порами (Кунце, Минних, по Шовену, 1953). При помощи антеннальных органов муравьи способны отличать чистую воду от подслащенной или ощущать в ней примесь кислоты или хинина (A. Schmidt, 1938). Порог чувствительности муравьев к сахарозе выше, чем у человека, и гораздо выше, чем у пчелы. Так, по данным Фриша (приведены до Шовену, 1953),
человек чувствует сахарозу при разбавлении молярного раствора в воде 1 : 80, пчела — 1:8 —1 : 16, Myrmica rubida I : 100, М. rubra — 1 : 150, a Lasius niger — 1 : 200.
О восприятии звука муравьями Шовен (1953) пишет следующее: «Муравьи реагируют на звук только тогда, когда они оказываются в центре стоячих волн, а не у вершины, как млекопитающие. У насекомых, не имеющих тимпанальных органов (муравьи), раздражением, вызывающим слуховые восприятия, по-видимому, является не изменение давления, а скорость движения молекул, максимальная в центре волн. Действительно, наблюдения показали, что некоторые волоски антенн начинают колебаться при помещении насекомого в центр волн, где амплитуда движения частиц уменьшена до 2 мк (Аутрум)». Вообще, по-видимому, звук для муравьев не играет существенной роли (Wilson, 1963b).
В разных местах на теле муравьев имеются небольшие участки, густо покрытые волосками, так называемые поля щетинок. Функциональное значение этих полей было недавно расшифровано Гюбером (R. Hubert, 1962). При помощи полей щетинок на усиках муравьи воспринимают движение воздуха. Другие поля являются рецепторами силы тяжести. При горизонтальном движении ориентацию осуществляют коксальные и брюшные поля, а при вертикальном — поля шеи, петиолюса, антенн и кокс. На F. polyctena показано путем последовательного исключения полей, что для правильной ориентации должна быть подвижной хотя бы одна из систем рецепторов.
Органами тактильного чувства (осязания) являются отстоящие волоски, расположенные на всем теле, и специальные органы усиков. При помощи этих же органов муравьи воспринимают сотрясения субстрата.
Половая система и ядовитые железы.

| 1 — половая система самца Formica sp.; 2 — половая система самки Formica rufa; 3-7 — последовательные стадии развития яйцевых трубочек малого лесного муравья; 3 — начальная стадия; 6 — полностью сформировавшиеся и функционирующие яйцевые трубочки; 7 — яицевые трубочки старой особи с резорбировавшимися яйцами. |
Половой аппарат самцов состоит из парных семенников, парных семенных протоков, которые затем сливаются в непарный семенной проток, открывающийся в эдеагус. На каждом из парных протоков перед их слиянием имеются семенные мешочки, служащие для хранения спермы. Семенники состоят из нескольких лопастей. У F. sanguinea, по данным Адлерца (по Wheeler, 1910), каждый семенник состоит из 21 лопасти. Половой аппарат самок состоит из большого количества яйцевых трубочек, открывающихся в парные яйцеводы, которые, сливаясь, образуют непарный яйцевод.
Семеприемник служит для хранения спермы, которая у муравьев сохраняется в течение всей жизни самок, так как оплодотворение у них однократное. Семеприемник снабжен специальной парной железой и открывается протоком в непарный яйцевод.
Развитие яиц до оплодотворения проходит в яйцевых трубочках. У самок F. rufa s. 1. их 45, у F. rufibarbis s. 1.— 18—20. У рабочих особей Formica также имеются яйцевые трубочки, но их значительно меньше. Так, у F. sanguinea их 3—6, у F. pratensis—2—6, у F. rufa (s. 1.) — 4—10 (Donisthorpe, 1915). Как показали последние исследования (Otto, 1958а и др ) У F. polyctena молодые рабочие имеют развитые функционирующие яичники (рис. 10, 3-6), а у старых особей яйца резорбируются.

1 — ядовитая железа: а — железистые отростки, б — секреторные трубочки, в — резервуар;
2 — железа Дюфура.
Из желез, принадлежащих половому аппарату , но изменивших свои функции, следует отметить ядовитую железу и железы Дюфура. У жалящих перепончатокрылых эти железы несут функцию ядовитых желез. Ядовитая железа имеет кислый экскрет, а дюфуровы — щелочной. У разных видов перепончатокрылых роль этих желез различна. У медоносной пчелы и шмелей, например, основное значение приобретают дюфуровы железы, a Formica являются крайним вариантом развития кислой ядовитой железы.
Ядовитая железа Formica состоит из большого мускульного резервуара служащего для хранения яда, и дорзальной железистой части. Железы представляют собой трубочки, которые одним концом открываются в центре резервуара, а с другого конца образуют парные железистые отростки Стенки трубочек состоят из полигональных клеток, каждая из которых имеет канал, начинающийся в цитоплазме и открывающийся в полость трубочки. В расправленном виде железы достигают 20 см (Wneeler, 1910).
Все представители подсемейства Formicinae не имеют жала и при защите используют челюсти и выбрызгивают экскрет ядовитой железы, причем в зависимости от преобладания того или иного способа защиты железа может быть развита по-разному (Stumper, 1952). Formica s. str. способны, сокращая мышцы резервуара, выбрасывать струю яда на расстояние около 20 см.
Состав яда Formica s. str. изучался многими авторами (Stumper, 1950, 1959а, b, 1960; Osman, Brander, 1961 и др.): 61-65% яда составляет муравьиная кислота (НСООН). Других кислот яд не содержит. 1.17-l.85% яда составляет сухое вещество, растворимое в ацетоне, в котором содержится 19,85% NH3 У зимующих или 4,83% NH3 У летних рабочих 15-17% аминокислот. Фосфатов в яде муравьев нет (Osman, Brander, 1961) Около 75% сухого вещества яда составляет пахучее вещество, по-видимому, тершшоид (Stumper, 1959a, b). Он образуется в железах Дюфура Штумпер (Stumper, 1959a, b) высказывает предположение, что это вещество является следовым феромоном, однако это предположе ние бездоказательно.
Количество муравьиной кислоты зависит от веса муравьев (Stumper, 1951). Через три недели истраченное содержимое резервуара восстанавливается (Sauerlander, 1961). Процесс образования кислоты в организме неизвестен, и на этот счет высказывается несколько гипотез (обзор — О Rourke, 1950b). Яд муравьев обладает инсектицидными антибиотическим действием. Инсектицидное действие оказывает только муравьиная кислота (Osman, Kloft, 1961). На лягушек она действует как нервный яд и влияет главным образом на работу сердца и дыхание (Цитович, Смирнов, 1915). Антибиотическое действие связано с другими компонентами яда (Sauerlander, 1961), возможно, с терпиноидным экскретом дюфуровых желез (btumper, 1959b).
В Германии в 1942 г. были произведены исследования дезинсекционного действия муравьиной кислоты (Hase, 1942). Кусочки тканей со всеми стадиями развития вшей помещались в муравейники рыжих лесных муравьев. Муравьи полностью очищали ткань за 6-24 час. В парах кислоты вши гибли через несколько часов, однако яйца оставались живыми.
Источник: Длусский Г.М. «Муравьи рода Formica»
Источник