
- Опыты Георгия Шапошникова по искусственной эволюции
- Биологические основы опытов
- Методика исследования
- Совместимость с новыми хозяевами
- Адаптация к новым хозяевам
- Dysaphis anthrisci chaerophyllina
- Dysaphis anthrisci majkopica
- Интенсивность естественного отбора
- Репродуктивная изоляция
- Жизненный цикл тлей рода Dysaphis
- Опыты по скрещиванию
- Интерпретация результатов
- 4.7. Тли Шапошникова
Опыты Георгия Шапошникова по искусственной эволюции

Опыты Георгия Шапошникова по искусственной эволюции — серия опытов, проведённых в конце 1950-х — начале 1960-х годов советским биологом Георгием Христофоровичем Шапошниковым, в процессе которых проводилась смена кормовых растений у различных видов тлей. Во время опытов впервые наблюдалась репродуктивная изоляция использованных в эксперименте особей от исходной популяции, что свидетельствует об образовании нового вида.
Биологические основы опытов
Большинство тлей чрезвычайно специфичны к выбору хозяина (кормового растения) и обитают на растениях только одного рода и даже одного вида. По большей части это определяется их пищевой специализацией, хотя иногда имеет значение строение ног и хоботка, приспособленные к определённому типу поверхности. Тем не менее, среди тлей время от времени происходит смена кормового растения, сопровождаемая долгим периодом пищевой адаптации. Г. Х. Шапошников стал одним из первых биологов, который экспериментально изучил этот процесс.
В 1940-х и 1950-х годах пищевую адаптацию тлей исследовали И. В. Кожанчиков, Е. С. Смирнов и З. Ф. Чувахина, однако они не довели опыты до логического завершения, проследив лишь небольшое количество поколений.
Методика исследования
Шапошников исследовал адаптацию при смене кормового растения для тлей подрода Dysaphis CB, s. str. Опыты проводились в 1954—1957 годах на хуторе Шунтук в районе Майкопа и в 1958 году в Ленинграде.
Первичными кормовыми растениями для всех видов тлей были яблони Malus orientalis Uglitz. и Malus domestica Borkh. В то же время в районе Майкопа тли могут мигрировать, приобретая вторичного хозяина, указанного в таблице.
Миграции различных видов тлей в районе Майкопа
Несмотря на то, что в литературе описаны случаи миграции тлей на чужеродные растения, Г. Шапошниковым в районе Майкопа ни одного подобного случая не обнаружено.
В отличие от опытов Е. С. Смирнова и З. Ф. Чувахиной, которые применяли посуточное чередование старого и нового хозяина, Шапошников использовал поиск случайных благоприятных сочетаний паразита и кормового растения. Во всех опытах использовались чистые клоны, произведённые партеногенетически от одной самки. В каждом случае формировалась популяция из 10 самок-клонов, которым предоставлялся выбор из нескольких вторичных хозяев, растущих в одном вазоне. Растения брались с разных станций, отличных по почвенным и климатическим условиям.
Клоны, потомки которых лучше всего прижились на новом кормовом растении, были в качестве контрольной группы размножены на 2—3 старых хозяевах и 3—10 новых.
Совместимость с новыми хозяевами
Без адаптации участвовавшие в опытах насекомые были в разной степени совместимы с новыми кормовыми растениями. Во всех опытах Dysaphis anthrisci chaerophyllina могли питаться и размножаться на Chaerophyllum bulbosum, но только в 9 % опытов они питались и в 4 % размножались на Anthriscus. Dysaphis anthrisci majkopica вообще не размножалась и только в 6 % опытов могли питаться на Chaerophyllum maculatum.
Совместимость (%) тлей с новыми кормовыми растениями
Адаптация к новым хозяевам
Dysaphis anthrisci chaerophyllina
В опытах с Dysaphis anthrisci chaerophyllina тли для формирования контрольной группы были пересажены на Chaerophyllum maculatum (вторичный хозяин), где прижились и до осени 1958 года дали на этом растении 47 поколений. Из контрольной группы насекомые пересаживались на новое кормовое растение (Chaerophyllum bulbosum), где также прижились, хотя до 8-го поколения их жизнедеятельность была неустойчивой из-за резких колебаний смертности.
В некоторых поколениях тли пересаживались с Chaerophyllum bulbosum обратно на вторичного хозяина Chaerophyllum maculatum, где нормально приживались.
Таким образом, тли Dysaphis anthrisci chaerophyllina в течение 4 поколений на новом хозяине (Chaerophyllum bulbosum) вполне освоили его, однако даже после 8 поколений не потеряли способность жить на старом хозяине (Chaerophyllum maculatum).
Dysaphis anthrisci majkopica
Пересадка Dysaphis anthrisci majkopica на Chaerophyllum maculatum происходила более сложным путём, поскольку на этом растении Dysaphis anthrisci majkopica без адаптации не способна размножаться. Первоначально насекомые были пересажены на малопригодное для них кормовое растение Chaerophyllum bulbosum, которое выполняло роль посредника. Затем насекомые, прижившиеся на Chaerophyllum bulbosum, пересаживались на Chaerophyllum maculatum.
Для формирования контрольной группы тли были пересажены на Anthriscus (вторичный хозяин), где прижились и размножались до осени 1958 года. С Anthriscus насекомые были пересажены на Chaerophyllum bulbosum. До 7-го поколения наблюдались резкие колебания плодовитости и смертности, однако начиная с 8-го поколения жизнедеятельность стабилизировались. На Chaerophyllum bulbosum тли прожили 11 поколений, до окончания жизненного цикла этих однолетних растений.
Начиная с 3—5 поколений тли с Chaerophyllum bulbosum регулярно пересаживались на Anthriscus. Начиная с 8-го поколения они потеряли способность питаться и размножаться на старом кормовом растении.
Пересадки Dysaphis anthrisci majkopica с Chaerophyllum bulbosum на Chaerophyllum maculatum показали, что в 8—9 поколениях тли получили способность питаться, а с 10—11 поколений нормально размножаться на ранее совершенно непригодном кормовом растении. Тли 10—11 поколений с Chaerophyllum bulbosum были пересажены на Chaerophyllum maculatum, где более года нормально жили и размножались.
Все попытки перевести Dysaphis anthrisci majkopica на Chaerophyllum maculatum непосредственно с Anthriscus потерпели неудачу за исключением одного случая, когда в 1958 году одна из 30 личинок 48-го поколения прижилась на Chaerophyllum maculatum. Было отмечено, что тли, воспитывавшиеся в Ленинграде в условиях, далёких от естественных, лучше приживались на чужеродных растениях.
Таким образом, тли Dysaphis anthrisci majkopica после 8 поколений адаптации к малопригодному Chaerophyllum bulbosum потеряли способность жить на старом хозяине (Anthriscus), но приобрели способность жить на новом, ранее совершенно непригодном Chaerophyllum maculatum.
Интенсивность естественного отбора
Средняя смертность личинок тлей на первичном кормовом растении была невелика, хотя для Anthriscus была ниже, что объясняется более высоким содержанием небелкового азота. Во время периода адаптации к новому кормовому растению смертность увеличивалась в десятки раз и в некоторых случаях достигала 100 %. По окончании периода адаптации смертность снижалась к уровню, характерному для видов, изначально специализировавшихся на данном растении.
Таким образом, естественный отбор интенсифицируется в период адаптации, в привычных же условиях он действует очень слабо. Процесс эволюции представляет собой чередование кратковременных периодов быстрых изменений с периодами относительного покоя и стабилизации.
Средняя смертность личинок (%) при воспитании на разных растениях
У Dysaphis anthrisci chaerophyllina адаптация шла в направлении использования нового, но пригодного корма. В результате интенсивность отбора была невелика, соответственно невелика была и глубина адаптации: освоив новое кормовое растение, тли не потеряли способность жить на старом. Адаптации такого уровня до Шапошникова исследовали Смирнов и Чувахина на примере оранжерейной тли Neomyzus circumflexus и Кожанчиков на примере листоедов Gastroidea viridula и Galerucella lineola.
У Dysaphis anthrisci majkopica шёл процесс освоения нового, ранее малопригодного корма. Интенсивность естественного отбора была чрезвычайно высока. В результате произошла глубокая адаптация, в процессе которой тли потеряли способность жить на старом кормовом растении.
Репродуктивная изоляция
Жизненный цикл тлей рода Dysaphis
Тли рода Dysaphis двудомны. На своём первичном хозяине (яблоне) они зимуют в виде яиц. Весной из яйца появляется самка-основательница и производит крылатых мигрантов, которые перелетают на вторичного хозяина — зонтичное растение. До осени мигранты размножаются путём партеногенеза и живорождения. Осенью на вторичном кормовом растении появляются ремигранты — самцы и т. н. полоноски, которые возвращаются к первичному хозяину. Здесь полоноски рождают бескрылых номальных (амфигонных) самок, которые спариваются с самцами и откладывают в трещины коры зимующие яйца.
Опыты по скрещиванию
В опытах по скрещиванию участвовали три популяции, перечисленные в таблице:
В опытах по скрещиванию нормальные самки одной формы скрещивались с самцами другой формы. Скрещивание производилось в прозрачных изоляторах, накрытых сетчатой тканью (мельничным газом). Кладка производилась либо в трещины коры (если спаривание происходило на яблоне), либо на кусочки бумаги, подкладываемые в изоляторы. Яйца зимовали в холодильнике, и весной из жизнеспособных яиц выходили самки-основательницы. Жизнеспособные яйца заметно отличались по внешнему виду — они были чёрными и блестящими в отличие от матовых, не чёрной окраски и часто сморщенных нежизнеспособных яиц.
Результаты скрещиваний (в просторных изоляторах на яблонях)
Опыты по скрещиванию показали, что
- Между Dysaphis anthrisci majkopica (m) и Dysaphis anthrisci chaerophyllina (c) существует практически полная репродуктивная изоляция. При скрещивании m×c (самцы m × самки c) жизнеспособные яйца вообще не появляются, а при скрещивании c×m — в среднем 1 жизнеспособное яйцо на 100 самок.
- После адаптации Dysaphis anthrisci majkopica (n) на ранее непригодном для него кормовом растении Chaerophyllum maculatum возникает практически полная репродуктивная изоляция с исходными видом Dysaphis anthriscimajkopica (m). При скрещивании m×n жизнеспособные яйца вообще не появляются, а при скрещивании n×m — не более 4 жизнеспособных яиц на 100 самок.
- У насекомых Dysaphis anthrisci majkopica (n), адаптировавшихся к Chaerophyllum maculatum возникает репродуктивная совместимость с аборигенным для этого растения подвидом Dysaphis anthrisci chaerophyllina (c). Скрещивание n×с по числу жизнеспособных яиц значительно превышает скрещивания с×с, n×n и m×m, однако скрещивание c×n уступает по этому критерию трём последним, что свидетельствует об определённой остаточной несовместимости.
Интерпретация результатов
Опыты Шапошникова вызвали сенсацию в среде советских биологов, хотя и остались практически неизвестными за рубежом, поскольку были опубликованы в русскоязычной периодике. Образование нового вида в течение нескольких поколений и в сроки, измеряемые месяцами, было явлением беспрецедентным.
Сам Шапошников выдвинул три возможных объяснения этому феномену, и все три признал чрезвычайно маловероятными:
- Вид n появился в результате проявления рецессивных признаков изначально гетерозиготных особей вида m: это объяснение маловероятно, поскольку размножение тлей в период адаптации происходило партеногенетически без скрещивания разнополых особей.
- Вид n является длительной модификацией m: это объяснение Шапошников отвергает, поскольку превращение m в n оказалось необратимым.
- Вид n явился результатом повторения (ресинтеза) эволюционного пути, который ранее привёл m в с: это объяснение Шапошников находит неправдоподобным, поскольку оно противоречит представлениям о необратимости эволюции.
В результате Шапошников не находит объяснения беспрецедентным результатам своих опытов, высказав лишь предположение, что виды c и n возникли из вида m в результате раскрытия неких «эволюционных потенций», изначально присутствовавших у вида m. На основании этого Шапошников классифицирует m, n и с как полувиды (не полностью сформировавшиеся виды), несмотря на наличие у них всех формальных признаков самостоятельных видов.
По мнению известного палеоботаника С. В. Мейена, опыты Шапошникова поставили под сомнение одну из концепций эволюционной теории — отсутствие соматической индукции (наследования приобретённых признаков). По этой причине они официально замалчивались и со временем оказались преданы забвению.
А. П. Расницин полагает, что быстрые эволюционные изменения в опытах Шапошникова связаны с ужесточением отбора только по одному признаку (непривычное питание) с одновременным ослаблением его по всем остальным (хищники, паразиты, болезни, неблагоприятные погодные условия). Аналогичные изменения наблюдаются у популяций, заселяющих обособленные территории (удалённые острова, озёра), где отсутствует конкуренция со стороны других видов.
Источник
4.7. Тли Шапошникова
В этом исследовании Шапошников тоже вывел «новый вид» тлей (очень быстро, практически за один сезон), которые смогли жить на новом растении хозяине, ранее абсолютно непригодном для этих тлей… после того, как Шапошников «потренировал» этих тлей на другом, «переходном» хозяине, более подходящем для исходных тлей.
Самое удивительное в этих исследованиях было то, что всё происходило очень быстро. Шапошников пересаживал примерно по 10 тлей на новое растение, а потом отсаживал обратно, для «восстановления жизненных сил». Таким образом, тли «тренировались» выживать на новом растении, на котором они исходно не питались, и не размножались. И по прошествии очень небольшого числа поколений (не более восьми!) тли вдруг начинали и питаться, и размножаться на новом растении. Причем этот переход всегда носил скачкообразный характер – сначала выживаемость тлей на новом растении имела неустойчивый характер. Но после определённого, конкретного поколения, тли вдруг дружно приживались на новом растении.
Остановлюсь только на самом ярком получившемся результате. Вот графики приживаемости тли Dysaphis anthrisci majkopica на новых (для этого вида тлей) растениях Chaerophyllum bulbosum и Сh. maculatum (Рис. 17):
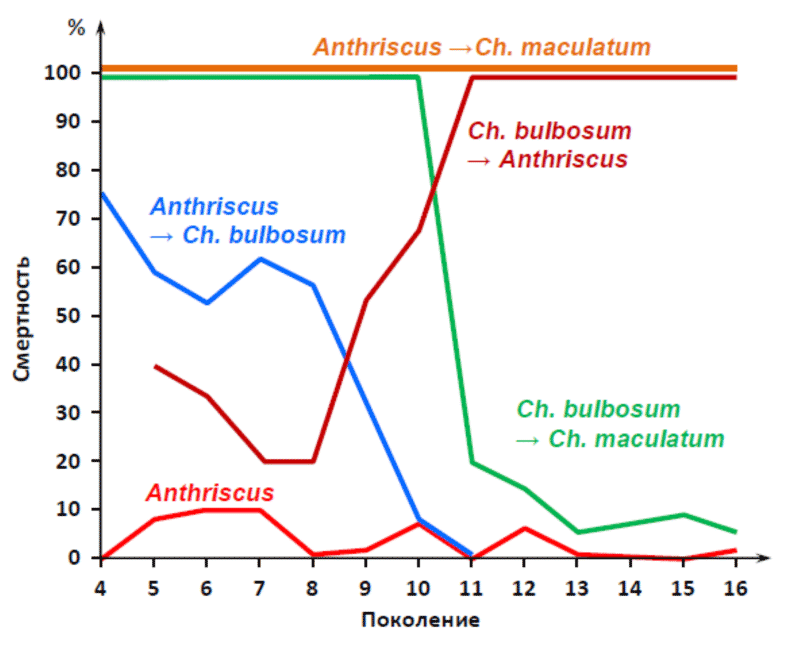
Рисунок 17. Графики, показывающие смертность и приживаемость тлей в ряду поколений на новых для них растениях в экспериментах Шапошникова.[36]
Из графика видно, что тли этого подвида:
1) Неплохо прижились на растении Anthriscus (красная линия внизу). Хорошо видно, что смертность тлей на этом растении – минимальна (это контрольная группа, которая никуда не пересаживалась).
2) При прямом пересаживании тлей с растения Anthriscus на растение Ch. maculatum – тли данного подвида не питались, не размножались и гарантированно (100 %) гибли (верхняя оранжевая линия).
3) Зато тли оказались способны «пересесть» c растения Anthriscus на Сh. bulbosum (синяя линия на графике). Причем до 8-го поколения наблюдались резкие колебания плодовитости и смертности, но после 8-го поколения – тли прижились на новом растении, и смертность резко упала.
4) Более того, тли, прижившиеся на новом растении (Ch. bulbosum), почему-то потеряли способность питаться на старом растении (Anthriscus). Из графика видно, что после всё того же восьмого (опять восьмого!) поколения – смертность этих «новых» тлей на старом растении резко повышается, достигнув к 11 поколению отметки в 100 % (темно-красная линия на графике, резко ползущая вверх).
5) В то же время, тли, прижившиеся на новом растении (Ch. bulbosum) – неожиданно приобрели способность питаться и размножаться на растении Ch. maculatum, которое изначально было вообще непригодным для исходных тлей данного подвида. Хорошо видно, что зеленая линия (смертности тлей) на графике крайне резко «падает» вниз на 10–11 поколениях. Что говорит о весьма синхронном «переключении» тлей. Это выяснилось после того, как тлей с Ch. bulbosum начали регулярно пересаживать на Ch. maculatum. В результате этих «тренировок», уже на 8–9 поколениях тли получили способность питаться на новом для них растении Ch. maculatum (причем дружно), а с 10–11 поколений уже нормально размножались на этом растении.
В итоге получилось, что тли необратимо «пересели» с одного растения на другое… всего за 11 поколений. Необратимо потому, что тли перестали питаться на старом растении. Более того, Шапошников провел скрещивания между полученными линиями тлей, и выяснил, что даже жизнеспособность оплодотворенных яиц между этими линиями упала чуть ли не до нуля, в то время как внутри линий – с жизнеспособностью яиц всё было хорошо (Шапошников, 1961). То есть, возникла, по сути, еще и репродуктивная изоляция.
Более того, при переходе тлей с одного растения на другое произошли еще и морфологические изменения тлей. Причем изменения весьма интересные – тли, перешедшие на новые растения… стали напоминать два других вида тлей (из этого же рода), всегда питавшихся на этих растениях. Например, проявилось сходство в строении хоботка и трубочек (Шапошников, 1965). То есть, получилась еще и определенная конвергенция форм (за 10 поколений).
Другой подвид тлей (Dysaphis anthrisci chaerophyllina) тоже повел себя весьма характерным образом:[37]
В опытах с A. chaerophyllina тли для формирования контрольной группы были пересажены на Ch. maculatum (вторичный хозяин), где прижились и до осени 1958 года дали на этом растении 47 поколений. Из контрольной группы насекомые пересаживались на новое кормовое растение (Ch. bulbosum), где также прижились, хотя до 8-го поколения их жизнедеятельность была неустойчивой из-за резких колебаний смертности.
И опять обратим внимание на назойливую цифру «8». Я затрудняюсь подробно интерпретировать эти результаты. Картина получается сложной, возможных вариантов объяснений много, и в них можно запутаться. Заострю внимание только на наиболее ярких деталях:
1. Мы опять наблюдаем очень быструю «эволюцию». Что довольно странно в рамках постулата о «случайных мутациях».
2. «Эволюция» успешно завершается каждый раз за 8 поколений, несмотря на очень малый биологический материал (около десятка «тренируемых» тлей). Что еще более странно для предположений о «случайных мутациях».
3. Наблюдается синхронность переходов тлей с одного пищевого объекта на другой. Постоянно фигурирует цифра – восемь (или около того). Это уже совсем противоречит предположению о «случайных мутациях».[38]
Возможно, все эти странности и можно как-то уместить в рамках постулируемого механизма «случайные мутации + естественный отбор». Но гораздо удобнее эти факты объясняются в рамках представлений о всё той же скрытой изменчивости (или даже о скрытых «переключателях» в геноме тлей), позволяющих им весьма быстро переключаться с одного субстрата на другой.
Интересно, что обсуждая полученные результаты, сам Шапошников в одних случаях пишет о «пищевых настройках» и «пищевых предпочтениях». Это когда пищевые изменения были обратимы (например, тли D. anthrisci chaerophyllina, перейдя на растение Ch. bulbosum, тем не менее, не потеряли возможность жить на исходном Ch. maculatum). А в других случаях Шапошников пишет о «пищевых адаптациях», в том числе, глубоких. Которые приводят к освоению нового растительного субстрата тлями, но с потерей способности питаться на прежнем субстрате. И далее рассуждает, являются ли «пищевые настройки», «пищевые предпочтения» и «пищевые адаптации» – проявлением неких скрытых потенций, то есть, возможностей, уже имевшихся у этих видов? Или же здесь имеет место изменение самих этих возможностей? Шапошников склоняется к тому, что в случае «пищевых настроек и предпочтений» реализуется первый вариант, а в случае «пищевых адаптаций» (во всяком случае, глубоких), реализуется второй вариант (Шапошников, 1961). То есть, Шапошников задаётся теми же вопросами, которыми сейчас задаёмся и мы. А что это такое продемонстрировали нам тли? Свои возможности по освоению новых видов кормовых растений, которые у них уже имелись в скрытом виде? Или же всё-таки настоящее изменение (эволюцию) своих пищевых адаптаций?
Более того, Шапошников приводит ряд других научных публикаций, где были зарегистрированы примеры таких же быстрых переходов насекомых фитофагов с одного растения на другое, которые тоже происходили за считанное число поколений. Например, установленные примеры с виноградной филлоксерой и т. п. (см. Шапошников, 1961). Получается, что такие быстрые переходы, возможно, достаточно широко распространены в природе. И является ли такой акт перехода насекомого-фитофага (с узкой специализацией) на новое (более-менее подходящее) растение – эволюционным актом? Или же эти насекомые «всегда так умели», то есть, такая способность (к быстрым пищевым переходам) исходна заложена в их геномах в виде «скрытых эволюционных потенций» – этот вопрос до сих пор висит в воздухе и «требует будущих исследований».
Источник