
- «Структурные основы регенерации поджелудочной железы»
- Кровавая драма на островке Лангерганса, или Как новооткрытая химера T- и В-клеток подстрекает аутоиммунный ответ при диабете I типа
- Конкурс «био/мол/текст»-2019
- Сравнительная морфология поджелудочной железы экспериментальных животных и человека
- Резюме
- Введение
- Развитие поджелудочной железы
- Анатомия
- Гистология
- Заключение
«Структурные основы регенерации поджелудочной железы»
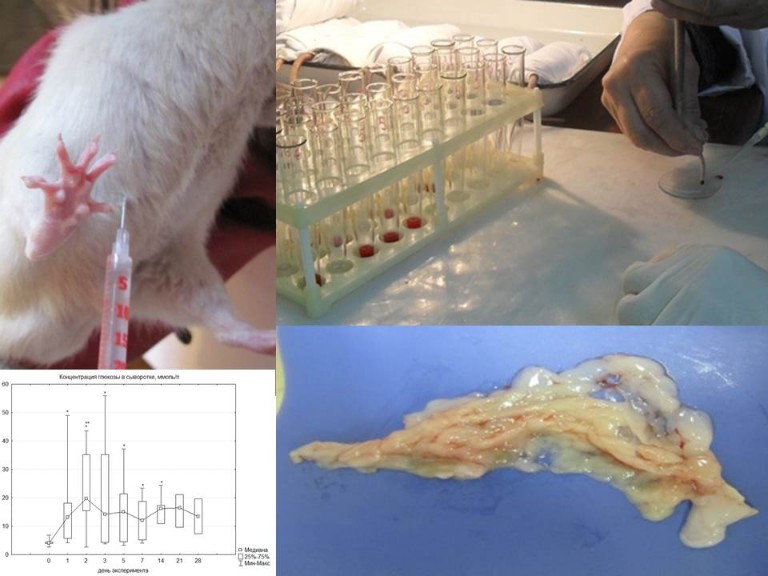
(рук. к.м.н., доц. М.С. Калигин, к.м.н., доц. С.Р. Абдулхаков)
Проект направлен на поиск и описание клеточных источников регенерации эндокринной части поджелудочной железы в ходе онтогенеза человека и в условиях экспериментально смоделированного сахарного диабета и медь-дефицита у крыс.
Результаты проекта:
— Получены данные экспрессии C-kit в ходе пренатального развития человека, которые позволяют отнести рецептор фактора стволовых клеток к маркерам коммитированых клеток-предшественниц инсулоцитов. C-kit-позитивные клетки-предшественницы инсулоцитов являются общими для α-и β-клеток островков Лангерганса и сохраняются в островках после рождения. То, что C-kit располагается на мембране клеток, делает его удобным инструментом для их выделения, изучения и разработки новых методов лечения сахарного диабета.
— Установлено, что в островках поджелудочной железы взрослого человека имеются C-kit-позитивные клетки, синтезирующие инсулин и глюкагон, поэтому можно говорить, что эти клетки находятся в состоянии дифференцировки в эндокриноциты. Пролиферации C-kit-позитивных клеток обнаружено не было, что может говорить о завершении процессов пролиферации этих клеток по крайней мере после 50 лет.
— В результате исследования экспериментального диабета уже на 1 сут. экспериментальной гипергликемии нами была обнаружена экспрессия C-kit в клетках островков поджелудочной железы, которая сохранялась на всех исследованных сроках до 28 суток. При этом клетки, имеющие на своей мембране C-kit, экпрессировали также инсулин и глюкагон. Таким образом, C-kit+ клетки, синтезирующие инсулин, могут способствовать коррекции нарушений углеводного обмена, возникающих при экспериментальном аллоксановом диабете.
— После частичной гепатэктомии в поджелудочной железе происходит активация С-kit+-прогениторных клеток и начинается их дифференцировка в α-клетки островков Лангерганса.
— Установлено, что уже через сутки экспериментальной гипергликемии обнаруживается экспрессия PCNA в клетках островков и ацинусов поджелудочной железы. Через 2 сут. также были обнаружены единичные PCNA-позитивные клетки с цитоплазматическим окрашиванием в интерстиции, около сосудов и протоков, показано, что данные клетки синтезируют инсулин. Таким образом, после повреждающего воздействия аллоксана на β-клетки островков при экспериментальном диабете I типа пролиферация возможна и в клетках островков, и в ацинарных клетках. Кроме того, результаты исследования подтверждают возможность пролиферации β-инсулоцитов или их предшественников.
— Установлено, что в ходе онтогенеза ПЖ человека первые MafA+ клетки появляются в островках на сроке 12,5 недель гестации, то есть позже, чем инсулин+ клетки (11,5 недель гестации) и глюкагон+ клетки (8,5 недель гестации). В эпителии протоков ПЖ MafA+ клеток не обнаружено. В ходе онтогенеза ПЖ количество MafA+ клеток в островках увеличивается и достигает максимума у взрослого человека, экспрессия MafA коррелирует с экспрессией инсулина и маркирует β-клетки. Результаты нашего исследования не позволяют рассматривать MafA в качестве маркёра КП инсулоцитов. Исходя из этого, мы считаем, что наши результаты подтверждают гипотезу, которая относит MafA к маркёрам зрелых β-клеток.
— В результате проведённых исследований экспериментального диабета у крыс установлено максимальное количество MafA-позитивных клеток в островках поджелудочной железы в норме и их уменьшение в островках на всех сроках экспериментального диабета у крыс при увеличении количества инсулин-позитивных клеток к концу первой и третьей неделям экспериментального диабета. Также установлено отсутствие схожести динамик изменений MafA- и C-kit-позитивных клеток при экспериментальном диабете. Результаты нашего исследования не позволяют рассматривать MafA в качестве маркёра недифференцированных предшественников инсулоцитов. Исходя из этого, мы считаем, что наши результаты вносят дополнительный вклад в гипотезу, которая относит MafA к маркёрам зрелых β-клеток.
Источник
Кровавая драма на островке Лангерганса, или Как новооткрытая химера T- и В-клеток подстрекает аутоиммунный ответ при диабете I типа
Коварная «бирецепторная» (dual-receptor-expressing) химера и другие клетки иммунной системы рядом с островком Лангерганса поджелудочной железы
Авторы
Редакторы
Комикс на конкурс «био/мол/текст»: Сахарный диабет первого типа, также известный как инсулинозависимый диабет, развивается из-за разрушения бета-клеток поджелудочной железы, продуцирующих инсулин — гормон, регулирующий уровень глюкозы в крови человека. В своей статье мы расскажем о том, как недавно открытый лимфоцит, несущий рецепторы как Т-, так и В-клеток, ошибочно подстрекает Т-клетки к атаке на инсулинпродуцирующие клетки.

Конкурс «био/мол/текст»-2019
Эта работа опубликована в номинации «Наглядно о ненаглядном» конкурса «био/мол/текст»-2019.

Генеральный спонсор конкурса и партнер номинации «Сколтех» — Центр наук о жизни Сколтеха.
Спонсор конкурса — компания «Диаэм»: крупнейший поставщик оборудования, реагентов и расходных материалов для биологических исследований и производств.

Спонсором приза зрительских симпатий выступила компания BioVitrum.
Давайте познакомимся с героями нашей статьи
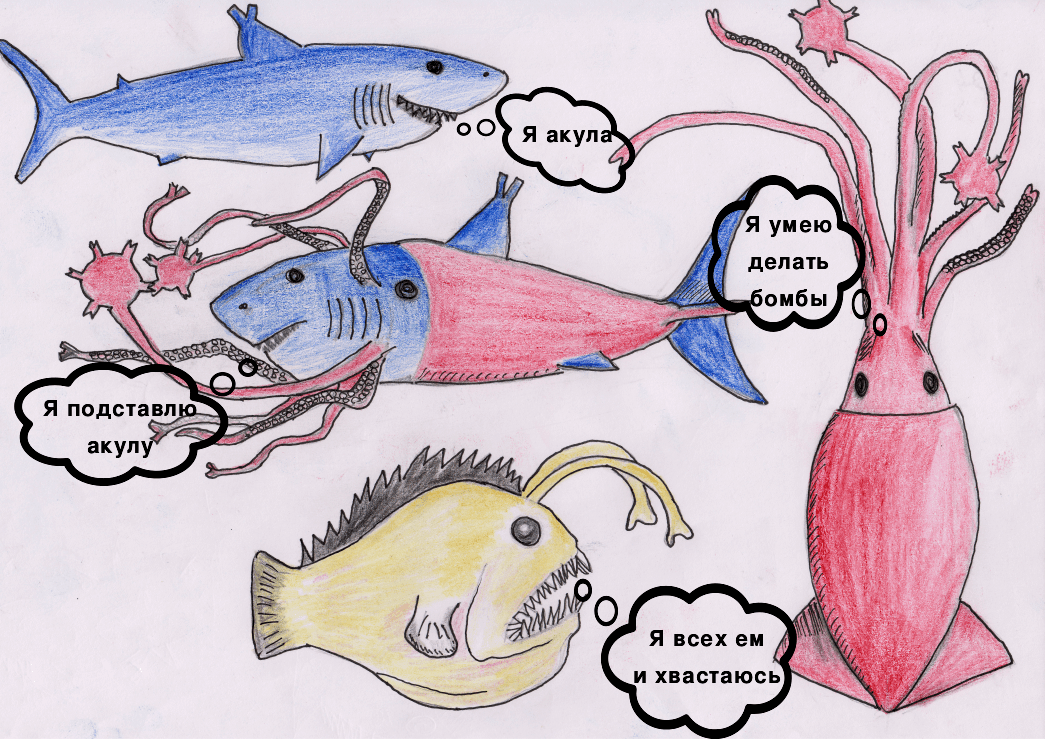
Акула — T-клетка.
Кальмар — В-клетка.
Удильщик — APC (антигенпрезентирующая клетка) с HLA I и HLA II (human leukocyte antigen).
Химера — DE-клетка (dual-receptor-expressing cell, «бирецепторная» клетка) с TCR (T-клеточным рецептором) и BCR (В-клеточным рецептором).
Для того чтобы разобраться, что же делает уникальный «бирецепторный» лимфоцит при диабете I типа (СД1), вкратце расскажем о том, как работает иммунная система.

Перед нами грозный хищник тканей организма — антигенпрезентирующая клетка (APC). Она поглощает фагоцитозом непрошенных гостей — патогенов, например, бактерии.

После переваривания она презентирует на специальном белке HLA II (human leucocyte antigen class II) часть белка патогена — антиген. Помимо HLA II существует HLA I. Об их значении и функциях подробнее рассказано в статье «Иммунитет: борьба с чужими и. своими» [1]. APC заплывает в лимфатический фолликул и привлекает туда наивные T-клетки, которые свободно циркулируют по организму.
В норме главная задача Т-лимфоцита — связывание антигена, который не встречается в организме, но попадает в него в течение патологических процессов. Разнообразие T-лимфоцитов очень велико. Оно обусловлено разнообразием Т-клеточных рецепторов, которое получается за счет рекомбинаций нескольких геномных участков и вставки случайных нуклеотидов в ген TCR [2]. Каждая из клеток немного отличается от остальных за счет гипермутабельности короткого фрагмента N-концевого домена рецептора и способна к узнаванию своего потенциального антигена. А значит, есть вероятность того, что APC встретит ту T-клетку, TCR которой будет связываться с антигеном, презентированным на HLA. Однако это также означает, что TCR может подойти и к пептиду, который есть у своих, нормальных клеток организма. Но нет. Такие Т-клетки обычно погибают в тимусе во время процесса, называемого негативной селекцией.
После связывания HLA II с TCR происходит пролиферация нужных T-клеток и выделение цитокинов, которые помогают им дифференцироваться. Для того чтобы эти T-клетки смогли связаться с HLA II, у них есть белок CD4, который «подтверждает» и стабилизирует связывание.

После дифференцировки CD4+ T-клетки выходят из фолликула и исполняют свою роль командира. Они выделяют цитокины, которые привлекают к месту инфекции другие клетки (базофилы, эозинофилы, тучные клетки и т.д.), и активируют макрофаги.
Тем временем наивные В-клетки (В-лимфоциты) ждут в фолликуле, когда приплывет антиген и они получат возможность принять участие в борьбе с патогеном.

Они действуют посредством поверхностных рецепторных молекул — В-клеточных рецепторов. В-клеточные рецепторы создаются с помощью высоко вариабельных генных перестроек. Наивная В-клетка ждет связывания своего BCR с антигеном. Она поглощает и представляет антиген на своем HLA II, и ждет встречи с CD4+ Т-клеткой, которая уже активировалась на тот же антиген. Прелесть рецепторов B-клеток (BCR) заключается в том, что благодаря случайной перестройке генов производится такое большое разнообразие рецепторов поверхности B-клеток, что практически любой чужеродный антиген, попадающий в организм, распознается ими.
T-лимфоцит помогает В-лимфоциту пролиферировать и начать продуцировать антитела, которые будут способны так же, как и В-клеточные рецепторы связывать антигены. Антитела связываются с антигенами на поверхности бактерий, а другим концом (Fc-концом) связываются с рецепторами на макрофагах (морская звезда на рисунке выше), после чего они фагоцитируют бактерию. Помимо этого, антитела на поверхности бактерий активируют систему комплемента, а также мешают бактерии прикрепляться к клеткам организма.
У 90% больных сахарным диабетом первого типа существует особый вариант HLA II (HLA-DQ8): он лучше связывается с антигеном, который несет относительный отрицательный заряд в определенных положениях (позициях 1 и 9). Инсулин считается аутоантигеном при СД1, то есть именно его распознают в качестве чужеродного агента иммунные клетки пациентов.

Обычно любой белок презентируется не целиком: клетки показывают только его наиболее иммуногенную часть. У инсулина такой частью являются аминокислоты 9–23 на В-цепи (B:9–23). Тем не менее презентация B:9–23 с помощью как нормального, так и мутантного (HLA-DQ8) HLA II для CD4+ T-клеток больных СД1 не приводит к их активации.
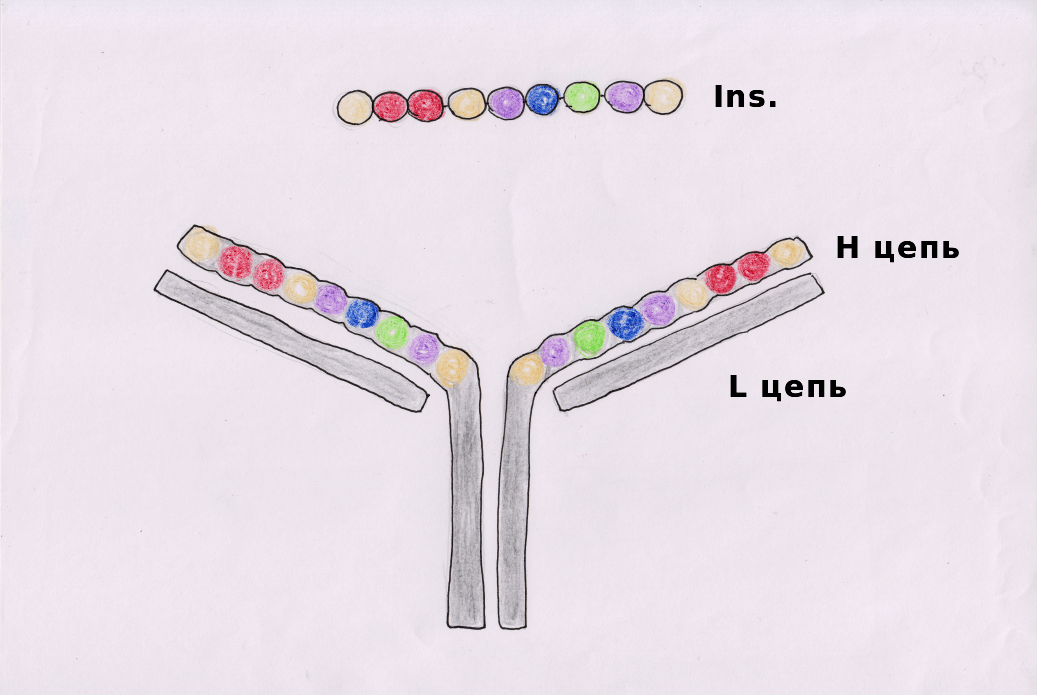
Недавно было открыто, что кровь людей, больных СД1, в сравнении со здоровой выборкой, оказывается обогащенной химерами Т- и В-клеток (Х-клетками) [3]. Антитела этих клеток имеют схожий с инсулином порядок зарядов аминокислотных остатков. Участки антител имеют лучшее сродство с инсулинспецифичными Т-клетками и активируют их, подстрекая развитие аутоиммунного ответа на инсулинпродуцирующие бета-клетки островков Лангерганса.

К сожалению, на данный момент неизвестно, какова природа возникновения химер Т- и В-клеток. Участие Х-клеток в развитии других заболеваний также остается загадкой.
Авторы предлагают проводить скрининг большего числа субъектов из группы риска и использовать данные об обогащении клонов Х-клеток в качестве прогностического признака. Также необходимо провести более качественный анализ RNA-seq, чтобы определить, являются ли Х-клетки отдельным новым типом клеток или присутствуют в организме в качестве субпопуляции одного из уже известных типов клеток.
Число людей, на которых было сделано это открытие, мало, но если результаты подтвердятся, то это станет захватывающим шагом к лучшему пониманию аутоиммунной природы сахарного диабета первого типа. Будет интересно увидеть будущие результаты этих исследователей.
Источник
Сравнительная морфология поджелудочной железы экспериментальных животных и человека
Я.И. Гущин, научный сотрудник, патоморфолог, В.В. Шедько, кандидат ветеринарных наук, научный сотрудник, патоморфолог, А.А. Мужикян, кандидат ветеринарных наук, старший научный сотрудник, руководитель лаборатории гистологии и патоморфологии, М.Н. Макарова, директор, В.Г. Макаров, доктор медицинских наук, профессор, зам. директора по науке НПО «Дом Фармации» НПО «Дом Фармации» Российская Федерация, 188663, Ленинградская обл., Всеволожский р-н, г.п. Кузьмоловский, ул. Заводская, д. 3, корп. 245 E-mail: guschin.ya@doclinika.ru
Резюме
Поджелудочная железа – непарный паренхиматозный орган относится к железам смешанной секреции, включает в себя экзокринную и эндокринную части, участвует в пищеварении и регуляции углеводного, белкового и жирового обменов в тканях. Работа поджелудочной железы подчиняется тонкой нейро-гуморальной регуляции, ее патология приводит к серьезным последствиям. В доклинических исследованиях используются разные виды лабораторных животных, поджелудочная железа которых по своему строению и функции чрезвычайно схожа с человеческой. Однако есть и отличия, которые могут существенно влиять на результат эксперимента. Данное сравнительное исследование было направлено на определение схожести и различия в морфологии органа у человека и лабораторных животных. Согласно анализу данных литературы, выявлены основные черты сходства строения поджелудочной железы человека и лабораторных животных, как в анатомическом, так и гистологическом строении. Железа имеет одинаковые источник развития и закономерности органо- и гистогенеза. Выраженно отличается анатомическое строение железы: у человека – компактного типа, у рассматриваемых животных – скорее брыжеечного типа. Хотя мнения авторов по данному вопросу отличаются. Несмотря на разнообразие формы у каждого вида, в железе выделяют головку, тело и хвост. Количество протоков у разных животных также вызывает споры исследователей. В литературе можно найти описание нескольких вариантов протоковой системы у животных. Гистологическое строение железы также однотипно. Экзокринная часть не имеет существенных особенностей у разных видов. А в строении островкового аппарата можно выделить видовые различия в качественном и количественном составе инсулоцитов в островках Лангерганса. Данная статья может быть полезна не только фармакологам и токсикологам, работающим в области доклинических исследований, но и гистологам, физиологам и патологам, изучающих патологические процессы и механизмы функционирования поджелудочной железы.
Введение
Поджелудочная железа (pancreas) – непарный паренхиматозный орган, расположенный позади желудка, преимущественно ретроперитонеально (только передняя ее поверхность покрыта брюшиной). Она относится к железам смешанной секреции, поскольку включает в себя экзокринную и эндокринную части. В экзокринной части продуцируется панкреатический сок, богатый пищеварительными ферментами (трипсин, липаза, амилаза и пр.), а в эндокринной части вырабатываются различные гормоны (инсулин, глюкагон, соматостатин, (вазоактивный интестинальный полипептид – ВИП), панкреатический полипептид). Железа участвует в пищеварении, вырабатывая пищеварительные ферменты, поступающие в двенадцатиперстную кишку и осуществляющие гидролиз практически всех составных частей пищи, поступающей в организм [1]. Деятельность органа чрезвычайно важна для регуляции углеводного (энергетического), белкового и жирового обменов в тканях животных и человека [2–6]. Работа поджелудочной железы подчиняется тонкой нейрогуморальной регуляции, ее патология приводит к серьезным последствиям.
В доклинических исследованиях используются разные виды лабораторных животных, поджелудочная железа которых по своему строению и функции чрезвычайно схожа с человеческой. Однако есть и отличия, которые могут существенно влиять на результат эксперимента. Задача данного сравнительного исследования – выявить схожесть и различия в морфологии органа у человека и лабораторных животных. Такие знания могут быть полезны не только фармакологам и токсикологам, работающим в области доклинических исследований, но и для гистологов, физиологов и патологов, изучающих патологические процессы и механизмы функционирования поджелудочной железы.
Развитие поджелудочной железы
О развитии поджелудочной железы, как человека, так и лабораторных животных (преимущественно грызунов и, особенно, крыс), написано много работ, однако в них встречается число противоречий.
Установлено, что по источнику развития и закономерностям органо- и гистогенеза поджелудочная железа человека и белой крысы схожи [7, 8].
Закладка железы начинается у человека в конце 3-й начале 4-й недели эмбрионального развития, а у крыс – на 10–11-е сутки. Она из энтодермы и мезодермы и имеет несколько зачатков. У человека выделяют 2 зачатка – вентральный и дорзальный [2, 8], в то время как у животных выделяют 3 зачатка – 2 вентральных и дорзальный [7]. Однако есть авторы, которые выделяют у крыс только 2 зачатка [8].
Вентральный зачаток поджелудочной железы представляет собой выпячивание скопления энтодермы зародышевой средней кишки в толщу мезенхимы между листками вентральной брыжейки. Дорсальный зачаток поджелудочной железы, зарождающийся раньше, располагается каудальнее вентрального зачатка, является выпячиванием печеночного дивертикула и первичной кишечной трубки в толщу мезенхимы дорсальной брыжейки. Оба зачатка сближаются и сливаются (у человека на 6–7-й неделе [7] и на 2-й неделе – у крыс [8]), формируя единую закладку, которая располагается в толще мезенхимы дорсальной брыжейки. Затем происходит канализация (тубуляция) эпителиальных тяжей: они приобретают форму трубочек, стенки которых представлены одним слоем цилиндрического эпителия; их количество постепенно нарастает. На их боковых стенках и концах появляются выпячивания в виде почек – зачатки концевых отделов – ацинусов, стенка которых образована одним слоем цилиндрического (призматического) эпителия. Плотно прилежащие друг к другу концевые отделы, сгруппированные в скопления, формируют первичные дольки, отграниченные тонкими прослойками рыхлой соединительной ткани.
Примерно на 3-м месяце внутриутробного развития у человека [2, 8] и на 15–16-е сутки – у крыс [8] происходит формирование экзокринной части железы за счет дифференцировки островковых клеток предшественниц, клеток эпителия протоков, преобразования ацинарных клеток и пролиферации дифференцированных эндокринных клеток в островках [9]. Образованные островки располагаются в толще долек и, как правило, тесно контактируют с концевыми отделами или выводными протоками. Во 2-й половине пренатального периода поджелудочная железа имеет отчетливое дольчатое строение. После рождения гистогенез поджелудочной железы не завершается – по периферии органа можно обнаружить эпителиальные трубочки с выпячиваниями, в которых продолжается активное деление клеток и формирование ткани железы [10], что связано с изменением условий функционирования организма.
Анатомия
Форма поджелудочной железы у человека и животных крайне разнообразна. Выделяют несколько типов строения: компактный, рассеянный или брыжеечный и промежуточный, сочетающий оба варианта [11]. У человека железа представляет собой компактный орган [2, 11, 12]. Большинство авторов сходятся во мнении, что поджелудочная железа крысы, мыши, кролика, морской свинки и хомяка классифицируется по брыжеечному типу. При таком типе ткани поджелудочной железы диффузно распределены в брыжейке двенадцатиперстной кишки, поперечной ободочной кишки и большого сальника, прилегающего к желудку и селезенке [3, 5, 13–17]. Так, у крысы, а в еще большей степени – у морской свинки, поджелудочная железа отличается своей рыхлостью, изогнутостью и разветвленностью [5, 15, 18, 19], так как состоит из отдельных маленьких гроздевидных долек, связанных рыхлой соединительной тканью в одно древовидно разветвляющееся образование [3, 14].
Однако Minoru Tsuchitani (2016) классифицирует поджелудочную железу крысы и мыши как промежуточный тип, поскольку селезеночная часть у указанных животных относительно компактна, но часть диспергирована в брыжейке. Он относит поджелудочную железу хомяков к 1-му – концентрированному типу. Поджелудочная железа кролика относительно мала. Она располагается в брыжейке между поперечной ободочной кишкой, желудком и двенадцатиперстной кишкой [20, 21] и состоит из отдельных частей неправильной формы, которые размещены в перитонеальной складке [22]. Дольки имеют, как правило, яйцевидную форму, 2–3 мм в диаметре и около 5 мм толщиной [23]. По мнению большинства авторов, такое строение – рассеянный, брыжеечный тип [11, 24]. Однако В.Н. Жеденов (1957) допускает существование как рассеянного, так и концентрированного типа, при котором ткань собрана компактно и напоминает единый орган [25]. У морских свинок форма поджелудочной железы в брюшной полости имеет S-образную форму, схожую с таковой у человека, но в расправленном состоянии напоминает бабочку [15]. Головка поджелудочной железы почти полностью окружена двенадцатиперстной кишкой и отграничена от желудка луковицей двенадцатиперстной кишки [15].
Поджелудочная железа человека характеризуется компактным расположением и лежит в подкове двенадцатиперстной кишки, на уровне I–II поясничных позвонков и покрыта тонкой соединительнотканной капсулой. Позади нее находится позвоночник, аорта, нижняя полая и левая почечная вены, спереди располагается желудок [12]. У взрослого человека длина ее составляет 14–20 см, ширина 3–9 см, толщина 2–3 см, масса 70–100 г [34, 39]. В поджелудочной железе выделяют головку, тело и хвост.
Головка (caput pancreatis) охвачена двенадцатиперстной кишкой, задняя поверхность прилежит к правой почечной и воротной венам, передняя – к поперечной ободочной кишке. На границе с телом имеется вырезка (incisura pancreatis), в которой проходят верхняя брыжеечная артерия и вена.
Тело (corpus pancreatis) трапециевидной формы, и имеет заднюю поверхность и покрытые брюшиной нижнюю и переднюю поверхности. На передней поверхности выделяется выпуклость в сторону сальника – сальниковый бугор (tuber omentale), который обнаруживается только у человека и хомяка, но в области хвоста есть железы [26].
Хвост (cauda pancreatis) направляется влево и лежит рядом с селезенкой, левым надпочечником и левой почкой.
Несмотря на разнообразное строение, в поджелудочной железе лабораторных животных, как и в поджелудочной железе человека, условно выделяют 3 основные части – головка (дуоденальная часть), тело (пилорическая часть) и хвост (желудочно-селезеночная часть) [3, 16]. По данным Minoru Tsuchitani (2016), поджелудочная железа крысы разделена на 4 доли: желудочный, селезеночный, парабилиарный и двенадцатиперстный сегменты (именуемые также лопастями [11, 16, 27]), которые формируют 2 крайние формы строения и топографии – молоткообразную и трилистника, когда доли вдаются в смежные брыжейки.
Таким образом, расположение частей железы следующее: парабилиарный сегмент и двенадцатиперстный сегмент расположена на дуоденальной стороне, желудочно-селезеночный сегмент простирается до желудка и селезенки, концевая часть селезеночного сегмента заканчивается около ворот селезенки, каудальная часть двенадцатиперстного сегмента и дорсальная часть селезеночного сегмента часто соединены вместе и простираются до ободочной кишки (рис. 1) [11].
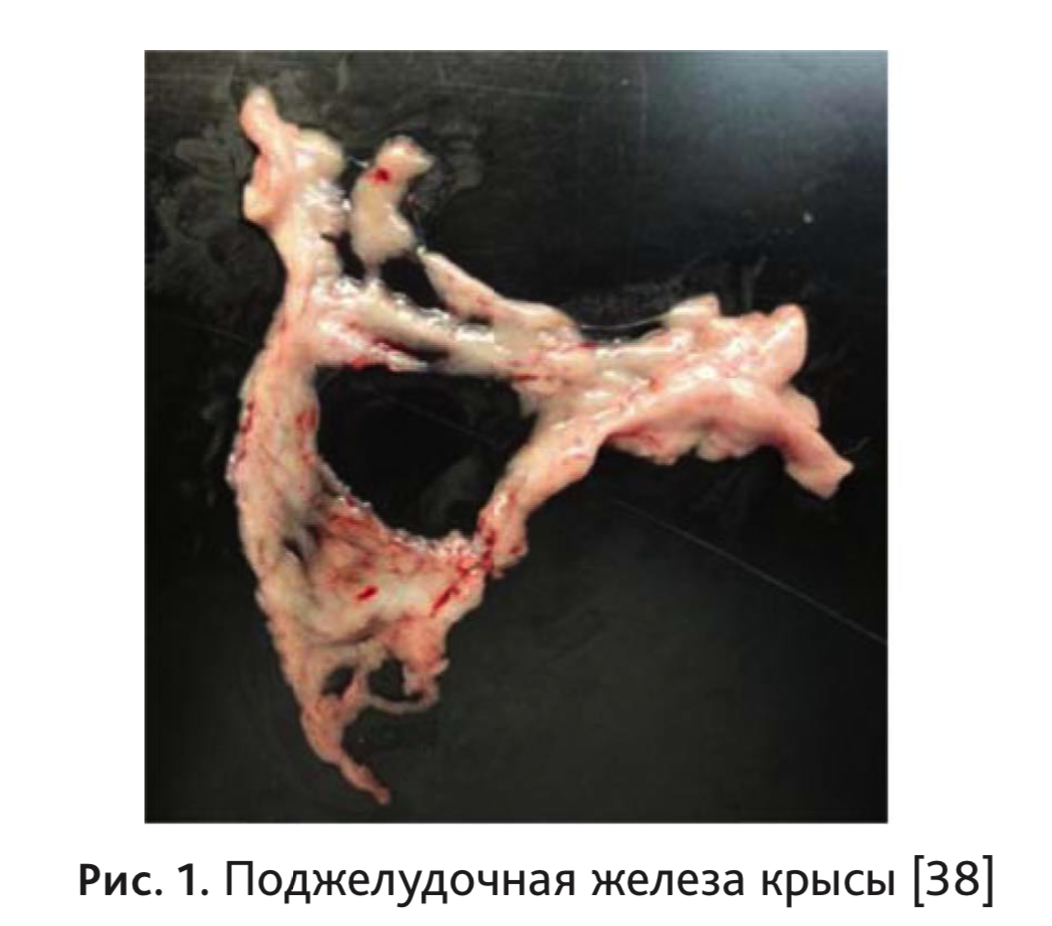
Протоки поджелудочной железы. У человека выделяют главный проток поджелудочной железы (ductus pancreaticus) или Вирсунгов проток, который принимает многочисленные ветви и, соединившись с общим желчным протоком, открывается общим большим сосочком двенадцатиперстной кишки (papilla duodeni major). Кроме того существует анатомически вариабельный добавочный или Санториниев проток (ductus pancreaticus accesorius), который начинается от головки поджелудочной железы и открывается в полость двенадцатиперстной кишки малым дуоденальным сосочком (papilla duodeni minor) [28].
На сегодняшний день мнения ученых о количестве выводных протоков поджелудочной железы у лабораторных животных расходятся. Так, по данным
М.Р. Бектемировой, особенностью строения поджелудочной железы крысы является наличие 2, очень тонких, выводных протоков, которые впадают либо в желчный проток, либо непосредственно в двенадцатиперстную кишку [3]. В литературе встречаются данные о наличии у крыс нескольких протоков поджелудочной железы, которые на различных уровнях впадают в общий желчный проток. По мнению ряда исследователей, у крыс, как и у мышей, общий желчный проток открывается в двенадцатиперстной кишке в большой сосочек двенадцатиперстной кишки. При этом отмечается, что у крыс, а также у морских свинок отсутствует малый сосочек [29, 30].
Кроме того, у мышей также есть 1 или несколько небольших дополнительных протоков поджелудочной железы, которые высвобождают содержимое непосредственно в двенадцатиперстную кишку, оканчиваясь при этом одним или несколькими малыми сосочками двенадцатиперстной кишки соответственно. K. Vashisht (2015) делает вывод, что анатомия выводных протоков поджелудочной железы у крыс и мышей несколько отличается.
Согласно другим исследованиям, у крыс и мышей междольковые протоки открываются в общий печеночный проток, который входит в двенадцатиперстную кишку, а также есть некоторые протоки, которые открываются непосредственно в двенадцатиперстную кишку [17], имеется и добавочный проток, который всегда впадает в кишку самостоятельно [31].
У кроликов главный панкреатический проток открывается около конца двенадцатиперстной кишки, вдали от входа желчных протоков, на расстоянии в 30–50 см от привратника [20, 21, 23]. Кроме того, есть добавочный проток поджелудочной железы, открывающийся самостоятельно в двенадцатиперстную кишку [20, 21]. Ученые сходятся во мнении, что добавочные протоки могут полноценно заменить собой главный в случае закупорки или экспериментальной перевязки главного [20, 21, 30]. Хотя по данным C.A. Johnson-Delaney (2006), существует 1 панкреатический проток, который дренирует обе доли и открывается на стыке поперечной и восходящей петли двенадцатиперстной кишки. При этом данный проток при эмбриональном развитии был добавочным, но впоследствии остался единственным, поскольку произошла облитерация терминальной части главного [32].
Кровоснабжение железы у человека и животных очень схожи [12, 25, 33–35]. Поджелудочная железа питается из бассейна чревного ствола ветвями печеночной артерии – желудочно-двенадцатиперстными артериями (arteriae pancreaticoduodenales), а также ветвями селезеночной артерии (arteria lienalis). Венозный отток происходит по одноименным венам в систему воротной вены.
Иннервация поджелудочной железы также однотипна. Она имеет симпатическую, парасимпатическую и автономную иннервацию. Парасимпатическая часть нервной системы регулирует секрецию гормонов посредством ветвей блуждающего нерва (nervus vagus) [12, 36]. Симпатические тела нервных клеток расположены в брыжеечных и брюшнополостных ганглиях.
Автономная нервная система, как у человека, так и у животных, распространена по всей железе в соединительнотканных перегородках и паренхиме [37, 38]. Интересно ее взаиморасположение с островковым аппаратом, поскольку вместе с инсулоцитами образуются нейроинсулярные комплексы 2 типов: I тип – смесь островковых и ганглиозных клеток, при типе II нервные волокна лежат рядом с инсулярными клетками [39].
Гистология
Снаружи железа покрыта тонкой соединительнотканной капсулой. Паренхима органа разделена на дольки (рис. 2), между которыми располагаются перегородки или септы – соединительнотканные тяжи с проходящими в них кровеносными сосудами, ганглиями и нервами, а также выводными протоками. Поджелудочная железа мыши, в отличие от других грызунов, имеет толстую капсулу из плотной волокнистой неоформленной соединительной ткани, сросшейся с висцеральным листком брюшины [40].
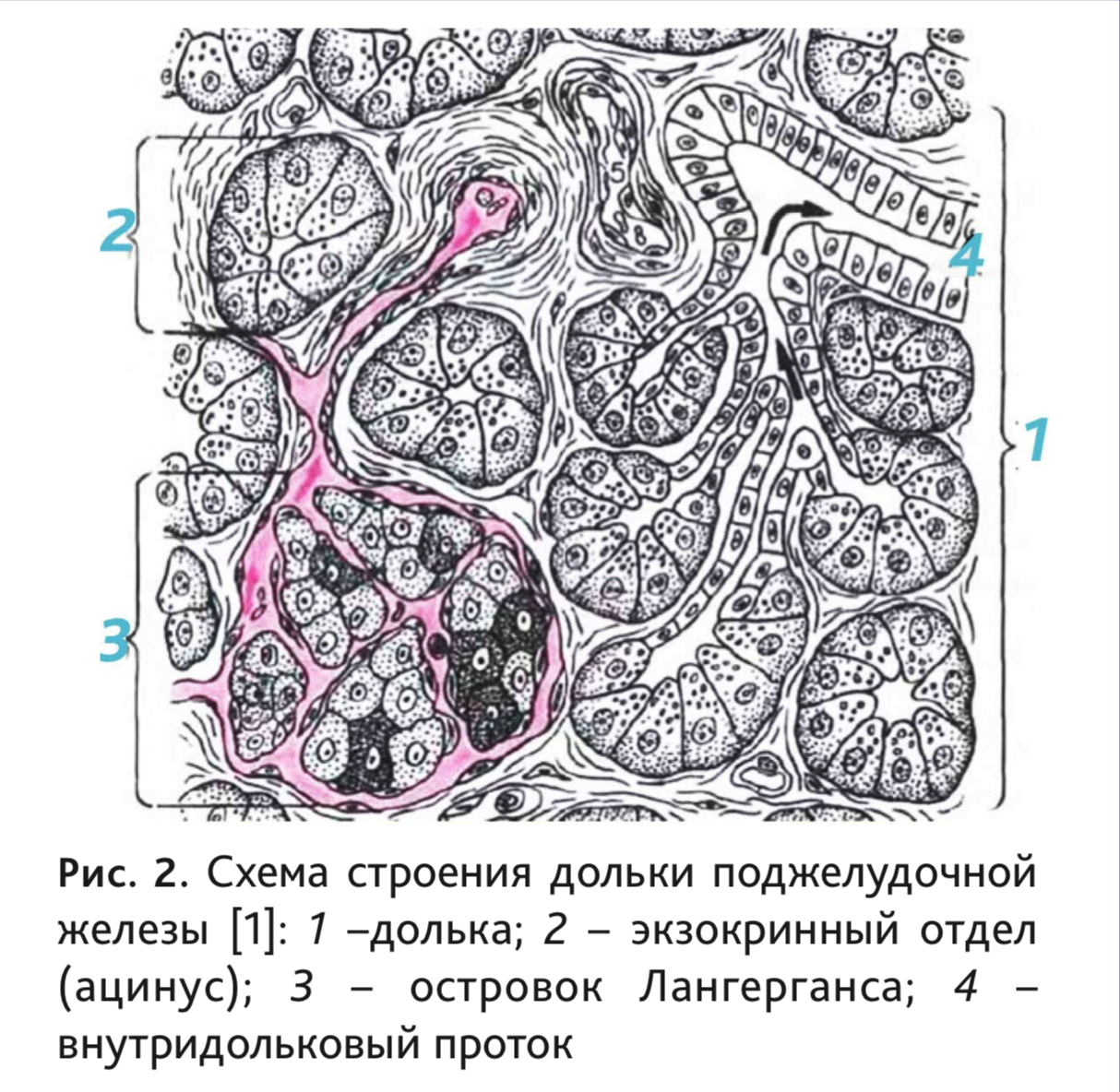
Экзокринная часть человека и рассматриваемых видов животных схожа по строению. Она состоит из структурно-функциональных единиц – ацинусов, а также системы протоков, включающих в себя вставочные, внутридольковые, междольковые протоки и общий панкреатический проток. Между ацину-
сами лежат ретикулярные волокна, капилляры, нервные волокна и ганг-
лии.
Ацинус состоит из 8–12 экзокринных панкреоцитов или ациноцитов, лежащих на базальной мембране и нескольких протоковых (центроацинарных) эпителиоцитов, образующих вставочный проток.
Ациноциты выполняют секреторную функцию и вырабатывают пищеварительные ферменты. Они имеют пирамидальную форму. Широкое основание лежит на базальной мембране. Узкая апикальная часть имеет микроворсинки, выступающие в просветы ацинуса. По восприимчивости красителей клетку можно разделить на верхушечную часть, или зимогенную зону и базальную часть, или гомогенную зону. Зимогенная зона окрашивается кислыми красителями и является оксифильной. В ней можно различить крупные секреторные гранулы, в которых находятся ферменты в неактивном состоянии, при этом гранулы имеют различную плотность, что определяет их зрелость. В гомогенной зоне осуществляется синтез ферментов, поэтому она пронизана гранулярной эндоплазматической сетью с большим количеством рибосом, что обусловливает базофильное окрашивание. В нижней части клеток располагается ядро, также содержащее 1–2 ядрышка. Надъядерную область занимает комплекс Гольджи, рядом с которым находится большое количество митохондрий.
Вставочный проток образован уплощенными клетками со светлой цитоплазмой, окружающей овальное ядро. Они могут примыкать к ацинарным клеткам с боку или проникать в центр ацинуса. Вставочный проток продолжается межацинозными протоками (ductus interacinosi), а затем переходит во внутридольковые протоки (ductus intralobulares). Стенки протоков выстланы однослойным кубическим эпителием, а вокруг них расположена рыхлая соединительная ткань с кровеносными капиллярами и нервами.
Внутридольковые протоки собираются в междольковые протоки (ductus interlobulares), которые лежат в междольковых перегородках и впадают в общий проток поджелудочной железы, который открывается в двенадцатиперстную кишку. Междольковые протоки и общий проток покрыты слизистой оболочкой, представленной высоким призматическим эпителием с небольшим количеством бокаловидных клеток и эндокриноцитов, которые вырабатывают панкреозимин и холецистокинин. Они лежат на собственной пластинке слизистой оболочки, в которой можно обнаружить мелкие слизистые железы [2, 11, 30, 36, 41].
Эндокринная часть представлена панкреатическими островками (insulae pancreaticae), или островками Лангерганса, которые лежат между ацинусами (рис. 3). У крыс клетки эндокринной ткани составляют менее 2% [11], у человека их количество не превышает 3% [2]. У человека они имеют округлую или овальную форму, но могут быть и лентовидными и звездчатыми [2]. Число, размеры, форма и микроскопическое строение островков Лангерганса не одинаковы у разных позвоночных [5], а их диаметр колеблется от 50 до 500 мкм [42].
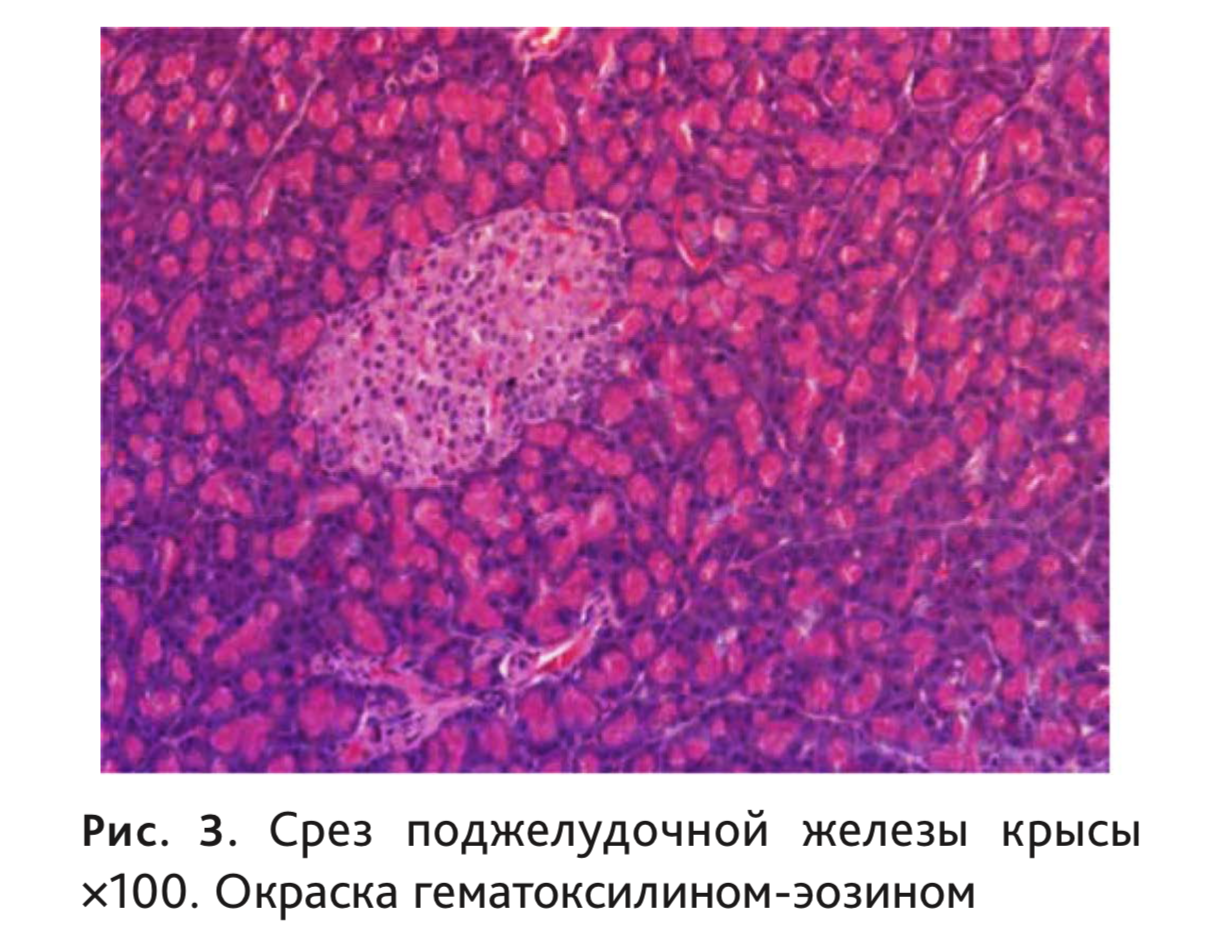
Состоят островки из инсулоцитов, вокруг которых лежат фенестрированные капилляры с перикапиллярным пространством, куда поступают синтезируемые гормоны. Капиллярная сеть островков сходна с клубочковой системой почки и имеет от 1 до 3 афферентных артериол. Плотность сосудов здесь в 5 раз выше, чем в экзокринной части.
Инсулоциты – небольшие клетки со светлой цитоплазмой, хорошо развитым аппаратом Гольджи, большим количеством митохондрий и секреторных гранул. Различают следующие виды клеток: базофильные (B-клетки), ацидофильные (A-клетки), дендритические (D-клетки) и РР-клетки. При светооптической микроскопии строение клеток у животных и человека схоже, но при электронно-микроскопическом исследовании можно обнаружить более выраженную видовую специфичность [11, 43] в распределении, размере и плотности гранул и органел. Так, например, у крыс секреторные гранулы В-клеток округлые с равномерной плотностью, а у морских свинок они могут быть полиморфными – дольчатыми, подковообразными различной плотности. У морских свинок средний диаметр А-гранул – 180 нм, В-гранул – 230 нм, а у кроликов размер А-гранул – 200 нм, В-гранул – 230 нм [43]. Кроме того, в панкреатических островках кроликов присутствуют фибриллярные структуры, не обнаруживающиеся в островковых клетках животных других видов [43]. Однако изучение ультраструктуры поджелудочной железы заслуживает отдельного рассмотрения. В данном обзоре подробно описать это невозможно.
A-клетки, на долю которых и у человека и у животных приходится до 25% всех клеток [2, 24, 44], имеют обычно неправильную форму и окрашиваются кислыми красителями в красный цвет. Их плотно расположенные в центре гранулы, отделенные от мембраны узким светлым ободком, содержат контринсулярный гормон – глюкагон. Характерной особенностью А-клеток является наличие в их цитоплазме многочисленных секреторных гранул, представляющих собой округлые тельца высокой электронной плотности, окруженные четко различимой гладкой мембраной. Между плотным содержимым секреторной гранулы и ее отграничивающей мембраной имеется узкая электроннопрозрачная зона (ореол). Электронноплотное «ядро» гранул обычно правильной округлой формы, его содержимое имеет мелкозернистый вид. Ядро А-клеток – округлое, расположено несколько эксцентрично. Митохондрии имеют удлиненную палочковидную форму и извилистые контуры. В цитоплазме некоторых А-клеток иногда имеется небольшое количество гранул гликоге-
на [42].
Основную массу клеток (у человека и у животных, по разным источникам, от 75 до 80% [2, 24, 44]) составляют B-клетки, которые окрашиваются базофильными красителями, а их гранулы содержат инсулин. Они обычно полигональной или неправильной формы, и являются наиболее крупными по размерам. Эндоплазматический ретикулум клеток развит хорошо, он образован короткими канальцами, а также небольшими вакуолями, на мембранах которых находятся связанные рибосомы. Имеется также значительное число свободных рибосом, расположенных поодиночке или небольшими группами между клеточными органеллами. Все это придает цитоплазме В-клеток более «темный» вид по сравнению с А-клетками. Удлиненные митохондрии многочисленны и сосредоточены в основном вблизи комплекса Гольджи, который развит хорошо и занимает большую площадь клетки. В-инсулоциты могут быть представлены «светлыми» клетками, в цитоплазме которых мало гранул, имеющих разные размеры и плотность. Они обычно распределены по цитоплазме или сконцентрированы в одном из полюсов. Органеллы представлены единичными цистернами гранулярного эндоплазматического ретикулума, элементами комплекса Гольджи и мелкими удлиненными митохондриями. «Темные» В-клетки содержат большое количество гранул разных размеров и интенсивности окраски, которые равномерно расположены в цитоплазме. В этих клетках органелл мало, в основном обнаружены мелкие митохондрии и несколько компактно расположенных мелких липидных включений. 3-й тип клеток, определяемый, как «промежуточный», имеет равномерное распределение среднего количества гранул, типичный набор элементов органелл, мелкие липидные включения, вакуоли. Среди 3 типов В-клеток можно встретить клетки с явными признаками деструкции: резкая вакуолизация цитоплазмы, иногда вытесняющие цитоплазму липидные включения [42].
D-клетки, количеством до 10% [2, 24, 44], в панкреатических островках встречаются довольно редко, они расположены поодиночке или небольшими группами по всему островку, имеют полигональную или звездчатую форму и длинные цитоплазматические отростки. D-клетки или непосредственно контактируют с капиллярами, или посылают к ним свои отростки. Характерной особенностью D-клеток является наличие в цитоплазме специфических секреторных гранул, размер которых значительно варьирует. Наибольшее скопление гранул наблюдается в цитоплазматических отростках и на сосудистом полюсе. Ядро D-клеток – крупное, имеет округлую или несколько вытянутую форму и содержит компактное ядрышко. Содержимое гранул – соматостатин, который обладает способностью тормозить секрецию инсулина и глюкагона, a также ограничивать всасывание глюкозы в кишечнике. Это влияние соматостатин может осуществлять паракринно, действуя как местный гормон [42].
Менее 1% полигональных мелких РР-клеток вырабатывают панкреатический полипептид. Их можно обнаружить по периферии островков, лежащих в области головки железы; единичные клетки определяются также в экзокринной части железы [2, 24, 44]. Они содержат в своей цитоплазме очень мелкие секреторные гранулы (80 нм). Последние располагаются обычно в 1 ряд вдоль плазматической мембраны [42].
Кроме того, существуют промежуточные или ациноостровковые клетки, которые располагаются среди экзокринной ткани вблизи панкреатических островков и характеризуются особо высокой активностью окислительных энзимов цикла Кребса – сукцинатдегидрогеназы, НАДН-дегидрогеназы, высокими показателями энергетического и белкового обмена. Эти клетки имеют ультраструктурные признаки, свойственные эндокринному и экзокринному типам клеток, одновременно синтезируя гормоны и пищеварительные ферменты. Особенность этих клеток с промежуточной экзокринно-эндокринной морфологией – наличие гранул – крупных зимогенных и мелких, характерных для островковых клеток, которые имеют разный гормональный профиль. Они содержат эндоплазматический ретикулум 2 типов – шероховатый и гладкий, митохондрии – крупные, свойственные экзокринным клеткам, и мелкие, типичные для островковых клеток. Экзокринные гранулы этих клеток располагаются в апикальных отделах, и их содержимое выделяется в просвет центрацинозных протоков. Эндокринные гранулы находятся в примыкающих к капиллярам базальных отделах, и их содержимое попадает непосредственно в кровь. По представлениям целого ряда исследователей, при определенных функциональных состояниях ациноостровковые клетки могут переходить в ацинарные или островковые [45].
Клеточный состав островков в поджелудочной железе имеет свои видовые отличия. У человека количество разных типов инсулоцитов распределяется как по островкам, так и внутри островка преимущественно равномерно, а у грызунов наблюдается разделение островков по преобладающим А- или В-клеткам, но чаще всего В-клетки, располагаясь в центре, формируют ядро, вокруг которого А-клетки образуют мантию с включением небольших групп и единичных клеток других типов (рис. 4). У морских свинок центральные В-клетки оттесняют другие клетки к периферии, но в центре можно выявить наличие групп D-клеток. Островковый аппарат кроликов отличается высокой гетерогенностью: островки могут состоять полностью из В-клеток или А-клеток, возможно также преобладание 1 типа клеток (чаще – В, но бывает даже D-тип), причем расположение их тоже разнообразно и вероятно, как центральное положение популяции инсулоцитов, так и смещение к периферии или полюсу островка [24, 44].
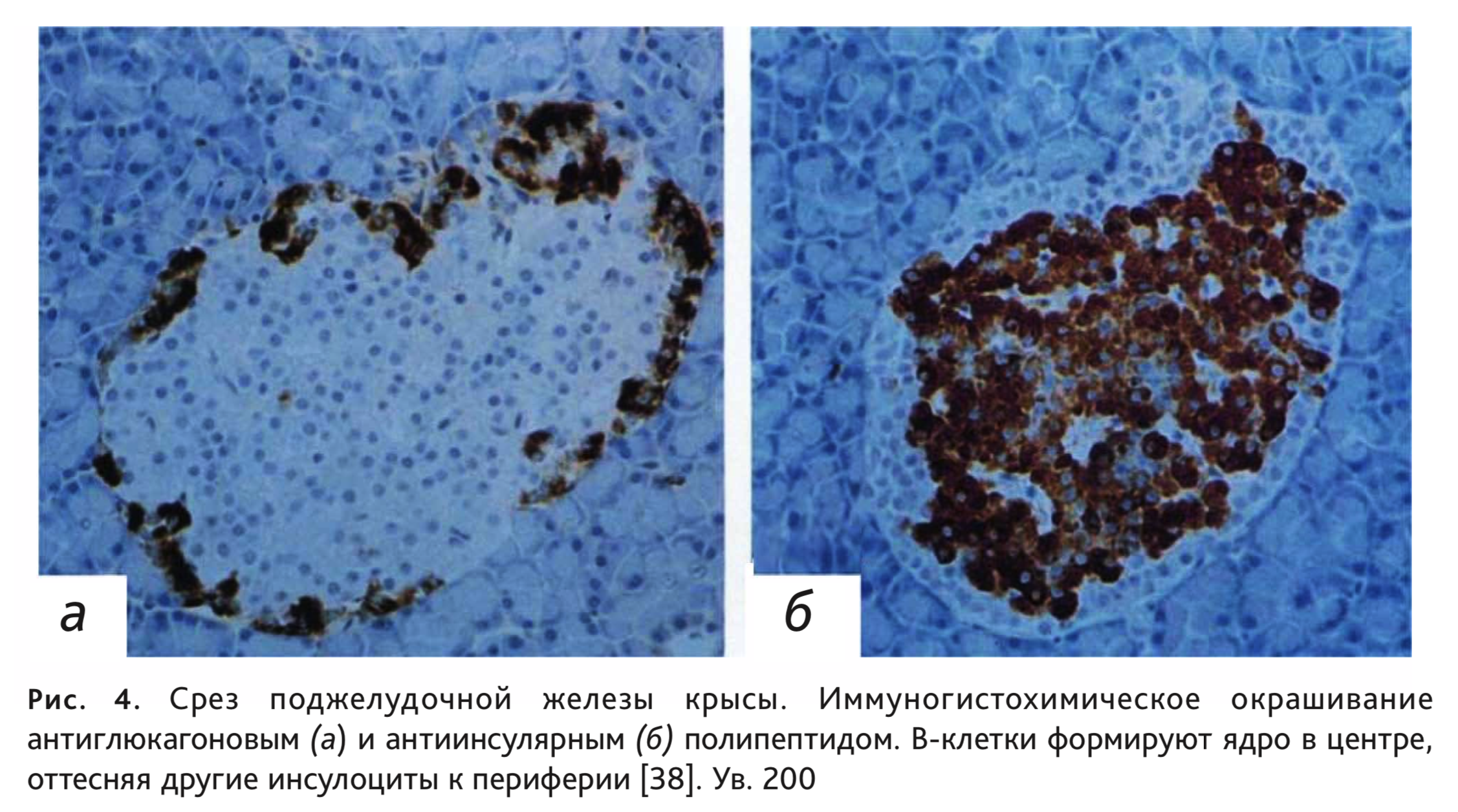
Кровоток в поджелудочной железе имеет свои особенности, он направлен от островков к ацинарной ткани. Структура микроциркуляторного русла представлена островково-экзокринной портальной системой. Прекапиллярные артериолы направляются к панкреатическим островкам, которые первыми получают артериальную кровь. Сразу же после вступления артериола распадается на капилляры, которые образуют своеобразный сосудистый клубочек. Кровь, оттекающая из островков и содержащая высокую концентрацию гормонов, попадает в капиллярную сеть экзокринной ткани поджелудочной железы, а не непосредственно в панкреатические вены, что обеспечивает возможность влияния этих гормонов на экзокринную паренхиму. Капиллярная внутриостровковая сеть имеет постоянное (векторное) направление кровотока от инсулин- к глюкагон- и далее – к соматостатин-секретирующим клеткам. Наличие инсулоацинарной портальной системы является одной из основ функционального взаимодействия экзокринной и эндокринной функций поджелудочной железы [23, 45, 46].
У человека капилляры имеют собственную мембрану, которая контактирует с мембраной инсулоцитов, формируя таким образом, перикапиллярное пространство. У мышей есть только одна – общая мембрана [36], но нет разделяющей базальной мембраны между панкреатическими островками и панкреатическими ацинусами. Между эндокринными и экзокринными клетками наблюдаются соединения в виде десмосом, интердигитаций, плотных и щелевидных контактов. На этом основании можно предположить, что клетки связаны друг с другом структурно и функционально [45].
Установлено, что регенеративные процессы тканей поджелудочной железы происходят не равномерно по всей ткани, а определёнными участками и в разное время, причем обновление в пределах каждой дольки происходит одновременно и в эндокринном, и в экзокринном отделах [9].
Самообновление в поджелудочной железе возможно при участии перибилиарных желез, располагающихся в стенке крупных желчных протоков или около них и открывающихся в их просвет. Установлено, что, помимо продукции муцина, в перибилиарных железах обнаружены ниши мультипотентных стволовых клеток, которые могут дифференцироваться в гепатоциты, холангиоциты и клетки островков поджелудочной железы. Перибилиарные железы внепеченочных желчных протоков описаны у человека, лабораторных животных (крыс, мышей), а также у большинства млекопитающих, но они не обнаружены они пока только у рептилий [47].
Заключение
Строение поджелудочной железы человека и лабораторных животных имеет много общего, как в анатомическом, так и гистологическом строении.
Источник развития и закономерности органо- и гистогенеза поджелудочной железы человека и животных схожи, но различаются сроками. Образовываясь из энто- и мезодермы, она имеет несколько зачатков: 2 – у человека и 3 – у грызунов. Однако вопрос органогенеза пока недостаточно изучен и характеризуется противоречивыми данными ряда авторов.
Наиболее яркое отличие имеет анатомическое строение железы, поскольку в зависимости от формы выделяют несколько типов строения органа: компактный, рассеянный (брыжеечный) и промежуточный. У человека поджелудочная железа компактная. Большинство авторов относит поджелудочную железу рассматриваемых животных к брыжеечному типу, но существуют мнения, что у хомяков строение органа компактное, у крыс и морских свинок смешанного типа, а у кроликов имеет место вариативность – возможен как брыжеечный тип, так и компактный. Несмотря на разнообразие формы у каждого вида в железе выделяют головку, тело и хвост.
Протоки поджелудочной железы также крайне разнообразны. У человека хорошо изучены варианты анатомического строения и преимущественно наблюдают 2 протока – главный и добавочный, которые открываются сосочками в двенадцатиперстную кишку. У животных, видимо, имеет место большая вариативность, поэтому описание протоков у разных авторов отличается. Так, у крыс выделяют протоки, которые впадают в желчный проток или сразу в двенадцатиперстную кишку, по другим данным, имеется всего 1 проток. У мышей описаны несколько дополнительных протоков открывающихся мелкими сосочками. Особенностью кроликов является значительное разобщение желчных протоков и протоков поджелудочной железы, но данные о количестве добавочных протоков весьма противоречивы.
Гистологическое строение железы также однотипно, значительную специфичность можно обнаружить только на ультрамикроскопическом уровне. Однако в данном обзоре это подробно не рассматривалось.
Экзокринная часть не имеет существенных различий. А в строении островкового аппарата можно выделить видовые различия в качественном и количественном составе инсулоцитов в островках Лангерганса. Так, например, у грызунов, в отличие от человека, присутствует разделение островков по преобладающим А- или В-клеткам, но чаще всего В-клетки, располагаясь в центре, формируя ядро, окруженное А-клетками и небольшими группами или единичными клетками других типов.
Источник