
- Химия, Биология, подготовка к ГИА и ЕГЭ
- Размножение партеногенезом
- Партеногенез у пресмыкающихся
- Механизм партеногенеза
- Происхождение однополых ящериц Различные аспекты эволюции партеногенеза у пресмыкающихся наиболее полно исследованы как раз в тех комплексах, у которых это явление и было впервые открыто, – у скальных и кнутохвостых ящериц. Из примерно 25 видов скальных ящериц 7 являются однополыми. В роде Cnemidophorus (45 видов) есть партеногенетические структуры как видового, так и под- и надвидового уровня. В целом не менее 30% популяций кнутохвостов состоят из одних лишь самок. Многочисленные исследования, проведенные на этих группах, позволили сделать несколько важных эволюционных заключений. Все партеногенетические виды образовались в результате гибридизации близкородственных двуполых видов. При этом в некоторых случаях процесс гибридизации происходил сложным образом и включал не два, а три родительских вида. Подобные события в комплексах скальных и кнутохвостых ящериц неудивительны. Ведь образующие их виды внешне и генетически очень сходны друг с другом, нередко обитают в одних и тех же местах, образуют сообщества и смешанные популяции. Появление среди них жизнеспособных гибридов, в том числе триплоидных и тетраплоидных, – явление обычное. Могут в таких комплексах образовываться и гибриды, способные к эндомитозу. А раз возникнув, такие организмы имеют много шансов сохраниться, поскольку им, чтобы оставить потомство, не нужны брачные партнеры. Партеногенетические формы оказались эволюционно юными. В отличие от других таксонов видового или подвидового уровня, их возраст оценивается не тысячами и миллионами лет, а всего лишь сот-нями. То, что среди известных однополых видов ящериц нет «долгожителей», позволяет предполагать эволюционную недолговечность партеногенетических форм. Однополые формы, в том числе и довольно широко распространенные, на всем протяжении своих ареалов представлены очень сходными – внешне и генетически – особями. Что, впрочем, не удивительно, если их происхождение действительно связано с немногими (или даже одним!) случаями гибридизации, а последующий эндомитоз не предполагает генетических изменений. Нужно отметить, однако, что существует и альтернативная гипотеза возникновения однополых ящериц. Согласно ей, партеногенетические самки могли спонтанно (как мутации) появляться в двуполых популяциях. Далее они давали начало однополым клонам, которые, в свою очередь, могли гибридизировать с близкими видами, образуя новые (генетически отличающиеся) однополые формы. Факультативный партеногенез у чешуйчатых пресмыкающихся Многие случаи партеногенеза, обнаруженные в последние годы у представителей разных семейств ящериц и змей, не связаны с существованием однополых популяций. Это – примеры факультативного партеногенеза. Как правило, они отмечены у животных, содержавшихся в неволе, в условиях изоляции от сородичей. Характерный пример – недавно опубликованное сообщение о партеногенезе у варана Varanus panoptes. Один немецкий террариумист приобрел пару молодых варанов для домашнего содержания. Оба животных оказались самками, агрессивно относившимися друг к другу. И в течение 5 лет их пришлось содержать в отдельных террариумах. Тем не менее обе самки отложили яйца. Любознательный владелец решил их на всякий случай положить в инкубатор. И оказался прав: из одного яйца вылупился детеныш-самец! И это несмотря на то, что его мать никогда в жизни не встречалась с самцами. То есть речь могла идти только о девственном размножении. В дальнейшем эту же самку ссаживали с самцом, и она приносила потомство уже обычным образом, после спаривания. Специалисты изучили генетическую природу неожиданного потомка, а также его младших братьев и сестер, появившихся на свет обычным путем, с помощью метода ДНК-фингерпринтинга, позволяющего достаточно просто оценивать различия в геномах отдельных особей для выявления их родства. Оказалось, что детеныши, родившиеся после спаривания, как и положено, имели часть локусов от матери и часть – от отца. А самый первый сын генетически не менее чем на 94% был идентичен своей матери. Тем не менее, он не является ее клоном – ведь они разнополые. Сравнение этого случая с известными цитогенетическими моделями партеногенеза не обнаружило полного соответствия ни одной из них. С уверенностью можно говорить лишь о том, что обнаруженный феномен – пример факультативного партеногенеза, требующий дальнейших исследований. Самцы у однополых видов ящериц Итак, изредка партеногенез может случаться в популяциях двуполых видов чешуйчатых пресмыкающихся. Однако возможна и зеркальная ситуация: в однополых популяциях спонтанно могут появляться самцы. Такой атавистический возврат к бисексуальности известен, например, у партеногенетических скальных ящериц и гекконов. У них регулярно находят мужские эмбрионы, которые, как правило, погибают на ранних стадиях развития. Но иногда они выживают, и родившиеся особи достигают зрелого возраста. Такие самцы партеногенетического происхождения часто обладают явными самцовыми вторичными половыми признаками (например, окраской) и проявляют свойственное самцам поведение (агрессивное, брачное). Но в то же время у них присутствуют и признаки гермафродитизма и к собственно размножению они обычно не способны. Эволюционные аспекты партеногенеза у чешуйчатых пресмыкающихся Эволюционный выигрыш однополых форм кажется очевидным. Потомство могут приносить все особи в популяции, а не только самки. То есть вдвое увеличивается репродуктивный потенциал популяции, а следовательно, возможности ее выживания и расселения. При этом экономятся энергетические ресурсы, которые обычно расходуются на поиск брачных партнеров, на брачные турниры, на ухаживание и спаривание. Кроме того, известно, что любая активность, связанная со спариванием, а также вся закрепляемая половым отбором внеш-няя «яркость» заметно повышают риск атаки хищников. То есть, теоретически, смертность бисексуальных видов от хищничества должна быть выше. Косвенное тому подтверждение – выраженно менее яркая и броская окраска партеногенетических видов скальных ящериц в сравнении с их бисексуальными сородичами. Еще одно явное преимущество однополых видов – практическое отсутствие нижнего порога численности жизнеспособной популяции. Известно, что у обычных видов снижение плотности популяции ниже определенного уровня приводит к практическому прекращению ее возобновления, – хотя еще есть и самки, и самцы, способные к размножению, им трудно разыскать себе партнера. А однополые формы могут успешно размножаться при любой плотности. Даже одна особь, случайно попавшая в новое место обитания, может стать основателем жизнеспособной колонии. Слабые стороны однополости менее очевидны, но, вероятно, перекрывают эти предполагаемые преимущества – в противном случае партеногенез был бы распространен в природе гораздо шире. Главный «недостаток» – низкое генетическое разнообразие в однополых популяциях, что снижает возможность реагировать на происходящие с течением времени изменения среды. Кроме того, детальные исследования показывают, что «очевидные» преимущества однополых видов в природе реализуются далеко не в полной мере. Так, их плодовитость в сравнении с бисексуальными родичами не так уже и высока. Например, у гекконов комплекса Heteronotia binoei плодовитость самок в партеногенетических популяциях примерно на 30% ниже, чем в обычных. У ряда однополых видов эмбриональная смертность оказывается выше, чем у близких двуполых форм. Способность к факультативному партеногенезу также имеет эволюционные преимущества. Она дает возможность оставить потомство в ситуациях резкого снижения численности или временного отсут-ствия контактов с другими особями. Подобное описано для водной бородавчатой змеи (Acrochordus arafurae). Бывает, что самки этого вида, оказавшиеся в мелких водоемах, не могут встретиться с самцами (поскольку по суше эти змеи перемещаются с трудом) и тогда начинают размножаться партеногенетически. С позиций эволюционного учения явление партеногенеза имеет и более общее значение, поскольку существенно расширяет возможности эволюционного формообразования. Появление партеногенетических групп изменяет систему репродуктивной изоляции близких видов. Спариваясь с самцами материнских или близкородственных видов, пар-теногенетические самки могут давать жизнеспособное потомство с принципиально новым набором генов – этот принцип лежит в основе концепции «сетчатой эволюции». Сходную роль в преодолении репродуктивной изоляции могут играть также самцы, появляющиеся изредка в однополых популяциях, если они будут давать потомство, спариваясь с самками других видов. Интересно, что принципиально иной способ размножения однополых рептилий позволяет лишь условно называть их ко-лонии «популяциями». Отсутствие генетического обмена между особями таких «популяций» делает неприменимым к ним и классическое определение биологического вида. По поводу таксономического положения таких форм между специалистами идет дискуссия. Например, исследователи скальных ящериц полагают, что партеногенез – дополнительная характеристика эволюционной обособленности. Поэтому те подвиды скальной ящерицы (Lacerta saxicola), у которых выявили однополость, были выделены в самостоятельные виды – Lacerta armeniaca, Lacerta rostombekovi. А вот герпетологи, изучавшие кнутохвостых ящериц, обнаружив гибридное происхождение однополых видов, напротив, сочли это аргументом против их генетической обособленности и стали считать их подвидами соответствующих родительских видов. Источник
- Факультативный партеногенез у чешуйчатых пресмыкающихся
- Самцы у однополых видов ящериц
- Эволюционные аспекты партеногенеза у чешуйчатых пресмыкающихся
Химия, Биология, подготовка к ГИА и ЕГЭ
В преддверии христианского праздника Пасхи я хотел бы рассмотреть тему, которая с научной точки зрения подходит к одному новозаветному чуду.
По приданию Пресвятая Дева Мария, без всякого оплодотворения, забеременела и родила Царя Иудейского Иисуса Христа – мессию, приход которого был предсказан в ветхом завете.
«Без оплодотворения? Не возможно!» – возразят некоторые. Но такое явление возможно. Деву Марию по-гречески называют «Агни Партене», переводится «Чистая Дева».
Вот от слова «партене» – дева, девственница — образован термин партеногенез.
Размножение партеногенезом

автор статьи — Саид Лутфуллин
Партеногенез – это процесс, при котором размножение происходит из неоплодотворенной яйцеклетки.
Но не следует путать это с бесполым размножением.
Размножение партеногенезом – это форма полового размножения, так как образуются женские гаметы .
Одними из первых партеногенез стали изучать шведский натуралист Шарль Бонне и немецкий зоолог Карл Зибольд.

Партеногенез делится на два вида: на мейотический и амейотический.
При амейотическом партеногенезе яйцеклетки остаются диплоидными, так как не претерпевают мейоза.
При мейотическом партеногенезе организм развивается либо из гаплоидной яйцеклетки, и сам является гаплоидным , либо яйцеклетка восстанавливает диплоидность и организм получается диплоидным .
Восстановление диплоидности может осуществляться по-разному: яйцеклетка может слиться с полярным тельцем (это похоже на копуляцию гамет) или может произойти эндомитоз.
Эндомитоз – процесс удвоения хромосом. Как при митозе, только не растворяется ядерная оболочка и не делится клетка.
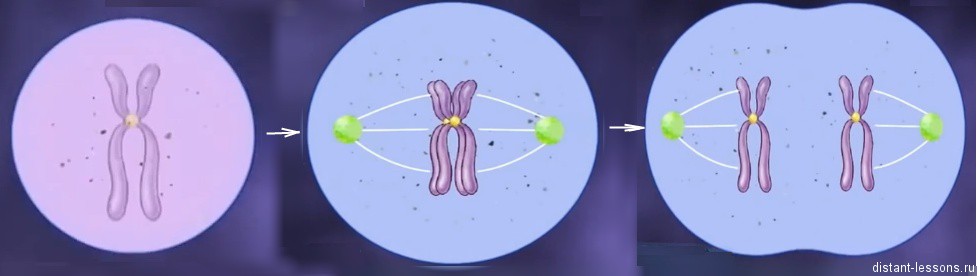
Какие же организмы могут размножаться партеногенезом?
Вот несколько классических примеров

Тли. Они таким образом быстро, без особых затрат увеличивают свою численность. Партеногенетически размножаются летом. В результате получаются только самки . Это своеобразная подготовка к неблагоприятным условиям, направленная на то, чтобы как можно больше особей выжили. С приближением осени на свет появляются другого типа гаметы, из которых могут появиться как самцы, так и самки. И насекомые начинают размножаться обыкновенным половым путем.
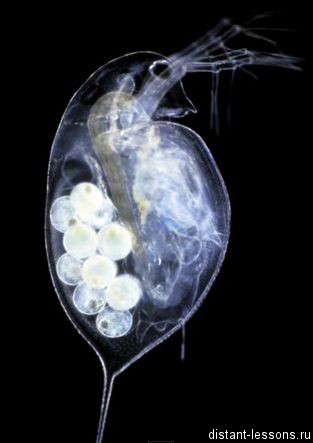
Дафнии (ракообразные). В течение лета размножаются амейотическим партеногенезом. Когда понижается температура водоема, сокращается световой день, появляются гаплоидные самцы. Популяция переходит к обыкновенному половому размножению.

Коловратки. Не удивляйтесь, если это название вам не знакомо, насколько мне известно, их нет в школьной программе. Если кратко: коловратки – это целый отдельный тип царства животные. Они многоклеточные организмы, но размеры их очень малы. У коловратки, так же как и тли и дафнии, размножаются партеногенезом в благоприятных условиях, а при наступлении неблагоприятных переходят к обыкновенному половому размножению. Есть даже некоторые виды типа коловратки, которые достигли «совершенства»: эти виды образованы только самками, которые размножаются партеногенезом. В таких случаях, когда партеногенез является единственным способом размножения, он называется облигатным. А когда наблюдается чередование партеногенеза и другого способа размножения партеногенез называется циклическим (как у дафний и тлей).

Пчелы. У пчел развитие яиц идет по двум схемам: некоторые оплодотворяются, некоторые нет. Из неоплодотворенных яиц (1n) развиваются самцы – трутни. Поэтому соматические клетки трутней гаплоидны ( об этом не нужно забывать, если вдруг в задаче по генетике попадется что-нибудь на эту тему ).
Из оплодотворенных яиц развиваются самки – рабочие пчелы либо матка. В таком случае, когда яйца могут развиваться и в результате оплодотворения, и партеногенетически, партеногенез называют факультативным.
Благодаря способности к факультативному партеногенезу у пчел осуществляется контроль численности особей каждой касты (рабочие, трутни).

Род Скальные ящерицы включает несколько видов, способных к партеногенезу. Перед мейозом в половых клетках этих ящериц происходит митотическое увеличение числа хромосом, поэтому после нормального цикла мейоза яйцеклетки получаются диплоидными и готовы образовать новый организм. Скальные ящерицы живут на камнях и иногда перебраться с одного на другой проблематично, в таких условиях как раз нужен партеногенез.
Обнаружен партеногенез у комодских варанов. Самки имеют половые хромосомы: ZW, а самцы: ZZ. Поэтому в результате партеногенеза должны получиться организмы: ZZ либо WW, но WW нежизнеспособны. Поэтому у комодских варанов в результате партеногенеза могут развиться только самцы.
Среди позвоночных размножение партеногенезом известно, помимо скальных ящериц и варана, у некоторых рыб и птиц.
Получается, непорочное зачатие – это всего лишь биологический процесс и никакого чуда нет?

Но не все так просто: у млекопитающих (включая человека) в естественных условиях размножение партеногенезом невозможно.
Так как у них наблюдается интересное явление – геномный импринтинг.
При геномном импринтинге в некоторых аллелях работают только гены, полученные от отца, или от матери. Поэтому организм не может развиться только из яйцеклетки, так как у него не будут работать некоторые гены. Так что место для чуда еще осталось.
Существует так же искусственный партеногенез. Путем различных внешних раздражителей можно заставить развиваться неоплодотворенную яйцеклетку. И уже получилось искусственно вызвать партеногенез у млекопитающих, однако получается это крайне редко.
Размножение партеногенезом у растений называют апомиксисом.
Источник
Партеногенез у пресмыкающихся
Наиболее заметное достижение отечественной герпетологии в минувшем веке, без сомнения, – открытие крупным российским зоологом Ильей Сергеевичем Даревским явления партеногенеза у ящериц.
Партеногенез – одна из форм полового размножения, при которой женские половые клетки развиваются без оплодотворения – широко распространен в царстве Флоры и среди различных беспозвоночных животных. Это было известно давно. Но для высокоорганизованных позвоночных партеногенез считался невозможным. до появления в 1958 г. сенсационной публикации И.С. Даревского в «Известиях Академии наук».
Начинающий тогда герпетолог, Даревский исследовал скальных ящериц Кавказа. Эти весьма обычные там ящерицы (относящиеся к обширному роду Lacerta из семейства настоящих ящериц) образуют сложный комплекс из множества внешне схожих видов, подвидов и рас, связанных сложными филогенетическими, географическими и экологическими отношениями. В настоящее время в этот комплекс включают около 25 видов, и некоторые систематики рассматривают его в качестве самостоятельного рода Даревския (Darevskia). Справедливости ради нужно сказать, что и более ранние исследователи отмечали отсутствие самцов в выборках из некоторых популяций скальных ящериц, но только Даревский понял, что эта странность не случайна, а представляет собой первое свидетельство существования феномена партеногенеза среди высших животных.

Уже через пару лет партеногенез был описан и в другом сложном комплексе ящериц – кнутохвостых, или кнемидофоров (род Cnemidophorus из североамериканского семейства Тейид, Teiidae).
После этих громких открытий внимание зоологов было привлечено к популяциям самых разных животных, в которых подозрительно преобладали самки или у которых самки приносили потомство без участия самцов. Как результат – целый каскад данных о партеногенезе в различных группах позвоночных. Существование партеногенетических популяций или появление потомства в результате однополого размножения было обнаружено у рыб, земноводных, птиц. К настоящему времени случаи однополого размножения не известны только среди млекопитающих, хотя и у представителей этого класса находят партеногенетические эмбрионы (которые, однако, погибают на ранних стадиях развития).
Но чешуйчатые пресмыкающиеся, и в первую очередь ящерицы, остаются самым интересным, важ-ным и перспективным объектом соответствующих научных исследований. Дело в том, что во всех остальных группах позвоночных партеногенез имеет факультативный характер, и только у ящериц (в 6 семействах, наиболее исследованные из которых – тейиды, настоящие ящерицы и гекконы) известны устойчивые популяции, состоящие из одних лишь самок. Партеногенез у чешуйчатых обнаружен также у игуан, хамелеонов, в семействах шипохвостов и ночных ящериц, а также у змей из семейств слепозмеек, удавовых, ужовых и ямкоголовых. Совсем недавно описаны случаи партеногенеза и у двух видов варанов, в том числе и у самой крупной из ныне живущих ящериц – комодского варана.
Механизм партеногенеза
Как известно, при обычном способе полового размножения после двух делений клетки в процессе мейоза образуются четыре гаплоидные гаметы, каждая из которых, соединяясь с гаметой особи противоположного пола, участвует в создании диплоидной зиготы. При партеногенезе же известно два основных механизма формирования генома потомства.
Одна форма однополого размножения – аутомиксис – также предполагает мейоз, однако образовавшиеся гаметы попарно соединяются друг с другом. При этом зигота может получить либо копии обеих материнских хромосом (центральное слияние), либо две копии одной из них (терминальное слияние).

Вторая форма – эндомитоз – с мейозом не связана. В этом случае происходит так называемое премейотическое удвоение хромосом с последующим делением клетки. Каждая дочерняя клетка получает точную копию генома матери (собственно, аналогично происходит и столь популярное сейчас клонирование). Именно этот механизм лежит в основе партеногенеза у однополых ящериц. У двуполых видов, у которых партеногенез отмечается лишь в отдельных случаях, он, очевидно, определяется аутомиксисом. Однако, поскольку случаи такие редки, их цитогенетическая природа изучена очень слабо.
Происхождение однополых ящериц
Различные аспекты эволюции партеногенеза у пресмыкающихся наиболее полно исследованы как раз в тех комплексах, у которых это явление и было впервые открыто, – у скальных и кнутохвостых ящериц. Из примерно 25 видов скальных ящериц 7 являются однополыми. В роде Cnemidophorus (45 видов) есть партеногенетические структуры как видового, так и под- и надвидового уровня. В целом не менее 30% популяций кнутохвостов состоят из одних лишь самок.
Многочисленные исследования, проведенные на этих группах, позволили сделать несколько важных эволюционных заключений.
Все партеногенетические виды образовались в результате гибридизации близкородственных двуполых видов. При этом в некоторых случаях процесс гибридизации происходил сложным образом и включал не два, а три родительских вида. Подобные события в комплексах скальных и кнутохвостых ящериц неудивительны. Ведь образующие их виды внешне и генетически очень сходны друг с другом, нередко обитают в одних и тех же местах, образуют сообщества и смешанные популяции. Появление среди них жизнеспособных гибридов, в том числе триплоидных и тетраплоидных, – явление обычное.
Могут в таких комплексах образовываться и гибриды, способные к эндомитозу. А раз возникнув, такие организмы имеют много шансов сохраниться, поскольку им, чтобы оставить потомство, не нужны брачные партнеры.
Партеногенетические формы оказались эволюционно юными. В отличие от других таксонов видового или подвидового уровня, их возраст оценивается не тысячами и миллионами лет, а всего лишь сот-нями. То, что среди известных однополых видов ящериц нет «долгожителей», позволяет предполагать эволюционную недолговечность партеногенетических форм.
Однополые формы, в том числе и довольно широко распространенные, на всем протяжении своих ареалов представлены очень сходными – внешне и генетически – особями. Что, впрочем, не удивительно, если их происхождение действительно связано с немногими (или даже одним!) случаями гибридизации, а последующий эндомитоз не предполагает генетических изменений.
Нужно отметить, однако, что существует и альтернативная гипотеза возникновения однополых ящериц. Согласно ей, партеногенетические самки могли спонтанно (как мутации) появляться в двуполых популяциях. Далее они давали начало однополым клонам, которые, в свою очередь, могли гибридизировать с близкими видами, образуя новые (генетически отличающиеся) однополые формы.
Факультативный партеногенез у чешуйчатых пресмыкающихся
Многие случаи партеногенеза, обнаруженные в последние годы у представителей разных семейств ящериц и змей, не связаны с существованием однополых популяций. Это – примеры факультативного партеногенеза. Как правило, они отмечены у животных, содержавшихся в неволе, в условиях изоляции от сородичей.
Характерный пример – недавно опубликованное сообщение о партеногенезе у варана Varanus panoptes. Один немецкий террариумист приобрел пару молодых варанов для домашнего содержания. Оба животных оказались самками, агрессивно относившимися друг к другу. И в течение 5 лет их пришлось содержать в отдельных террариумах. Тем не менее обе самки отложили яйца. Любознательный владелец решил их на всякий случай положить в инкубатор. И оказался прав: из одного яйца вылупился детеныш-самец! И это несмотря на то, что его мать никогда в жизни не встречалась с самцами. То есть речь могла идти только о девственном размножении.

В дальнейшем эту же самку ссаживали с самцом, и она приносила потомство уже обычным образом, после спаривания. Специалисты изучили генетическую природу неожиданного потомка, а также его младших братьев и сестер, появившихся на свет обычным путем, с помощью метода ДНК-фингерпринтинга, позволяющего достаточно просто оценивать различия в геномах отдельных особей для выявления их родства. Оказалось, что детеныши, родившиеся после спаривания, как и положено, имели часть локусов от матери и часть – от отца. А самый первый сын генетически не менее чем на 94% был идентичен своей матери. Тем не менее, он не является ее клоном – ведь они разнополые. Сравнение этого случая с известными цитогенетическими моделями партеногенеза не обнаружило полного соответствия ни одной из них. С уверенностью можно говорить лишь о том, что обнаруженный феномен – пример факультативного партеногенеза, требующий дальнейших исследований.
Самцы у однополых видов ящериц
Итак, изредка партеногенез может случаться в популяциях двуполых видов чешуйчатых пресмыкающихся. Однако возможна и зеркальная ситуация: в однополых популяциях спонтанно могут появляться самцы. Такой атавистический возврат к бисексуальности известен, например, у партеногенетических скальных ящериц и гекконов. У них регулярно находят мужские эмбрионы, которые, как правило, погибают на ранних стадиях развития. Но иногда они выживают, и родившиеся особи достигают зрелого возраста. Такие самцы партеногенетического происхождения часто обладают явными самцовыми вторичными половыми признаками (например, окраской) и проявляют свойственное самцам поведение (агрессивное, брачное). Но в то же время у них присутствуют и признаки гермафродитизма и к собственно размножению они обычно не способны.
Эволюционные аспекты партеногенеза у чешуйчатых пресмыкающихся
Эволюционный выигрыш однополых форм кажется очевидным. Потомство могут приносить все особи в популяции, а не только самки. То есть вдвое увеличивается репродуктивный потенциал популяции, а следовательно, возможности ее выживания и расселения. При этом экономятся энергетические ресурсы, которые обычно расходуются на поиск брачных партнеров, на брачные турниры, на ухаживание и спаривание. Кроме того, известно, что любая активность, связанная со спариванием, а также вся закрепляемая половым отбором внеш-няя «яркость» заметно повышают риск атаки хищников. То есть, теоретически, смертность бисексуальных видов от хищничества должна быть выше. Косвенное тому подтверждение – выраженно менее яркая и броская окраска партеногенетических видов скальных ящериц в сравнении с их бисексуальными сородичами.
Еще одно явное преимущество однополых видов – практическое отсутствие нижнего порога численности жизнеспособной популяции. Известно, что у обычных видов снижение плотности популяции ниже определенного уровня приводит к практическому прекращению ее возобновления, – хотя еще есть и самки, и самцы, способные к размножению, им трудно разыскать себе партнера. А однополые формы могут успешно размножаться при любой плотности. Даже одна особь, случайно попавшая в новое место обитания, может стать основателем жизнеспособной колонии.
Слабые стороны однополости менее очевидны, но, вероятно, перекрывают эти предполагаемые преимущества – в противном случае партеногенез был бы распространен в природе гораздо шире. Главный «недостаток» – низкое генетическое разнообразие в однополых популяциях, что снижает возможность реагировать на происходящие с течением времени изменения среды. Кроме того, детальные исследования показывают, что «очевидные» преимущества однополых видов в природе реализуются далеко не в полной мере. Так, их плодовитость в сравнении с бисексуальными родичами не так уже и высока. Например, у гекконов комплекса Heteronotia binoei плодовитость самок в партеногенетических популяциях примерно на 30% ниже, чем в обычных. У ряда однополых видов эмбриональная смертность оказывается выше, чем у близких двуполых форм.
Способность к факультативному партеногенезу также имеет эволюционные преимущества. Она дает возможность оставить потомство в ситуациях резкого снижения численности или временного отсут-ствия контактов с другими особями. Подобное описано для водной бородавчатой змеи (Acrochordus arafurae). Бывает, что самки этого вида, оказавшиеся в мелких водоемах, не могут встретиться с самцами (поскольку по суше эти змеи перемещаются с трудом) и тогда начинают размножаться партеногенетически.

С позиций эволюционного учения явление партеногенеза имеет и более общее значение, поскольку существенно расширяет возможности эволюционного формообразования. Появление партеногенетических групп изменяет систему репродуктивной изоляции близких видов. Спариваясь с самцами материнских или близкородственных видов, пар-теногенетические самки могут давать жизнеспособное потомство с принципиально новым набором генов – этот принцип лежит в основе концепции «сетчатой эволюции». Сходную роль в преодолении репродуктивной изоляции могут играть также самцы, появляющиеся изредка в однополых популяциях, если они будут давать потомство, спариваясь с самками других видов.
Интересно, что принципиально иной способ размножения однополых рептилий позволяет лишь условно называть их ко-лонии «популяциями». Отсутствие генетического обмена между особями таких «популяций» делает неприменимым к ним и классическое определение биологического вида. По поводу таксономического положения таких форм между специалистами идет дискуссия. Например, исследователи скальных ящериц полагают, что партеногенез – дополнительная характеристика эволюционной обособленности. Поэтому те подвиды скальной ящерицы (Lacerta saxicola), у которых выявили однополость, были выделены в самостоятельные виды – Lacerta armeniaca, Lacerta rostombekovi. А вот герпетологи, изучавшие кнутохвостых ящериц, обнаружив гибридное происхождение однополых видов, напротив, сочли это аргументом против их генетической обособленности и стали считать их подвидами соответствующих родительских видов.
Источник