
Тля — Aphid

Жоффруа , 1762 г.
- AphididaeLatreille, 1802 г.
- † BajsaphididaeHoman, Zyla & Wegierek, 2015 г.
- † CanadaphididaeRichards, 1966 г.
- † CretamyzidaeHeie, 1992
- † DrepanochaitophoridaeZhang & Hong, 1999
- † OviparosiphidaeShaposhnikov, 1979.
- † ParvaverrucosidaePoinar & Brown, 2006
- † SinaphididaeZhang, Zhang, Hou & Ma, 1989 г.
- incertae sedis
- † Palaeoforda tajmyrensisКононова, 1977
- † PenaphisLin, 1980 г.
- † Plioaphis subhercynicaHeie, 1968
- † SbenaphisScudder, 1890 г.
- † SunaphisHong & Wang, 1990 г.
- † КсилутянкаллисВан, 1991
- † ЮэфисВан, 1993
Тли маленькие сок -sucking насекомых и членов надсемейства Aphidoidea . Общие названия включают зеленую и мошку , хотя люди в пределах одного вида могут сильно различаться по цвету. В группу входят пушистые белые шерстистые тли . Типичный жизненный цикл включает нелетающих самок, которые рожают нимф женского пола, которые, возможно, уже беременны , что ученые называют телескопическим развитием, без участия самцов. Быстро созревая , самки обильно размножаются, поэтому количество этих насекомых быстро увеличивается. Крылатые самки могут развиваться позже в течение сезона, позволяя насекомым колонизировать новые растения. В регионах с умеренным климатом осенью наступает фаза полового размножения , когда насекомые часто зимуют в виде яиц .
Жизненный цикл некоторых видов включает чередование двух видов растений-хозяев, например, между однолетней культурой и древесным растением . Некоторые виды питаются только одним типом растений, в то время как другие являются универсальными, колонизируя многие группы растений. Описано около 5000 видов тлей, все они входят в семейство Aphididae . Около 400 из них встречаются на пищевых и волокнистых культурах , и многие из них являются серьезными вредителями для сельского и лесного хозяйства , а также доставляют неудобства садоводам . Так называемый Молочными муравьи имеют мутуалистических отношения с тлей, стремящимися их за пади , и защищая их от хищников .
Тля — одно из самых разрушительных насекомых-вредителей культурных растений в регионах с умеренным климатом. В дополнении к ослаблению растения, высасывая сок, они выступают в качестве векторов для вирусов растений и изуродовать декоративные растения с депозитами пади и последующим ростом копоти форм . Благодаря их способности быстро увеличиваться в численности путем бесполого размножения и телескопического развития, они представляют собой очень успешную группу организмов с экологической точки зрения.
Бороться с тлей непросто. Инсектициды не всегда дают надежные результаты, учитывая устойчивость к нескольким классам инсектицидов и тот факт, что тля часто питается нижней стороной листьев. В садовых масштабах довольно эффективны струи воды и мыльные распылители. Естественные враги — хищные божьи коровки , личинки журчалок , паразитические осы , личинки тлей-мошек , пауки-крабы , личинки златоглазок и энтомопатогенные грибы . Интегрированной борьбы с вредителями стратегии с использованием биологической борьбы с вредителями может работать, но трудно достичь только в закрытых помещениях , таких как теплицах .
СОДЕРЖАНИЕ
Распределение
Тля распространена по всему миру , но наиболее распространена в зонах умеренного климата . В отличие от многих таксонов , видовое разнообразие тлей в тропиках намного ниже, чем в зонах умеренного климата. Они могут мигрировать на большие расстояния, в основном за счет пассивного рассеивания ветрами. Крылатая тля также может подниматься днем на высоту до 600 м, куда ее разносит сильный ветер. Например, смородино-салатная тля Nasonovia ribisnigri , как полагают, распространилась из Новой Зеландии в Тасманию примерно в 2004 году благодаря восточным ветрам. Тля также распространяется при транспортировке зараженных растительных материалов людьми , что делает некоторые виды почти космополитичными по своему распространению.
Эволюция


История окаменелостей
Тля и близкородственные адельгиды и филлоксераны , вероятно, произошли от общего предка около 280 миллионов лет назад , в раннепермский период. Вероятно, они питались такими растениями, как Cordaitales или Cycadophyta . Из-за своего мягкого тела тля плохо окаменелость, и самая старая известная окаменелость относится к виду Triassoaphis cubitus из триасового периода . Однако иногда они застревают в экссудатах растений, которые превращаются в янтарь . В 1967 году, когда профессор Оле Хейе написал свою монографию « Исследования ископаемых тлей» , было описано около шестидесяти видов из триасового, юрского , мелового и в основном третичного периодов, а балтийский янтарь внес еще сорок видов. Общее количество видов было небольшим, но значительно увеличилось с появлением покрытосеменных 160 миллионов лет назад , поскольку это позволило тлям специализироваться, а видообразование тлей шло рука об руку с разнообразием цветковых растений. Самые ранние тли, вероятно, были многоядными , а монофагия развивалась позже. Была выдвинута гипотеза, что предки Adelgidae жили на хвойных деревьях, в то время как предки Aphididae питались соком Podocarpaceae или Araucariaceae, которые пережили вымирание в конце мелового периода. Такие органы, как роговицы, не появились до мелового периода. Одно исследование также предполагает, что предковые тли могли жить на коре покрытосеменных растений и что питание листьями может быть производным признаком . У Lachninae длинные части рта, которые подходят для жизни на коре, и было высказано предположение, что их предок в среднем меловом периоде питался корой покрытосеменных деревьев, а в позднем меловом периоде перешел на листья хвойных растений-хозяев. Phylloxeridae вполне может быть самым старым из до сих пор сохранившихся семейств, но их летопись окаменелостей ограничена палеофиллоксерами нижнего миоцена .
Таксономия
Реклассификации В конце 20-го века в полужесткокрылых уменьшило старый таксон «HOMOPTERA» на два подотряда: Sternorrhyncha (тли, белокрылки, весы , psyllids и т.д.) и цикадовые ( цикады , цикадки , treehoppers , дельфицидов и т.д.) с подотряда Heteroptera содержащая большую группу насекомых, известных как настоящие жуки . Инфраотряд Aphidomorpha внутри Sternorrhyncha варьируется в зависимости от очертания, при этом особенно трудно выделить несколько групп ископаемых, но они включают Adelgoidea, Aphidoidea и Phylloxeroidea. Некоторые авторы используют единое надсемейство Aphidoidea, в которое также включены Phylloxeridae и Adelgidae, в то время как у других есть Aphidoidea с сестринским надсемейством Phylloxeroidea, в которое входят Adelgidae и Phylloxeridae. Реклассификация в начале 21 века существенно изменила классификацию семейств Aphidoidea: некоторые старые семейства были понижены до уровня подсемейства ( например , Eriosomatidae ), а многие старые подсемейства были повышены до уровня семейства. Самые последние авторитетные классификации включают три надсемейства Adelgoidea, Phylloxeroidea и Aphidoidea. Aphidoidea включает одно большое семейство Aphididae, которое включает все
5000 существующих видов.
Филогения
Внешний
Тли, адельгиды и филлоксериды очень тесно связаны между собой и входят в подотряд Sternorrhyncha, жуков, сосущих растения. Они либо помещаются в суперсемейство насекомых Aphidoidea, либо в суперсемейство Phylloxeroidea, которое включает семейство Adelgidae и семейство Phylloxeridae. Как и тля, филлоксера питается корнями, листьями и побегами виноградных растений, но, в отличие от тли, не производит выделений пади или роговицы . Филлоксера ( Daktulosphaira vitifoliae ) — это насекомые, которые вызвали Великую французскую гниль вин , опустошившую европейское виноградарство в 19 веке. Точно так же адельгиды или шерстистая хвойная тля также питаются флоэмой растений и иногда описываются как тли, но более правильно классифицируются как тлейоподобные насекомые, потому что у них нет хвоста или роговиц.
Отношение к группам, особенно к ископаемым, сильно различается из-за трудностей в разрешении отношений. Большинство современных методов лечения включают три надсемейства, Adelogidea, Aphidoidea и Phylloxeroidea внутри отряда Aphidomorpha, а также несколько групп ископаемых, но другие методы лечения включают Aphidomorpha, содержащие Aphidoidea, из семейств Aphididae, Phylloxeridae и Adelgidae; или Aphidomorpha с двумя надсемействами, Aphidoidea и Phylloxeroidea, последнее содержит Phylloxeridae и Adelgidae. Филогенетическое дерево из Sternorrhyncha выводится из анализа малой субъединицы (18S) рибосомальной РНК .
Psylloidea (прыгающие тли и т. Д.)
Aleyrodoidea (белокрылки)
Phylloxeridae (филлоксераны)
Adelgidae (шерстистая хвойная тля)
Внутренний
Филогенетическое дерево, основанное на Papasotiropoulos 2013 и Kim 2011, с дополнениями из Ortiz-Rivas и Martinez-Torres 2009, показывает внутреннюю филогению Aphididae.
Было высказано предположение, что филогения групп тлей может быть выявлена путем изучения филогении их бактериальных эндосимбионтов , особенно облигатного эндосимбионта Buchnera . Результаты зависят от предположения, что симбионты передаются строго вертикально через поколения. Это предположение хорошо подтверждается доказательствами, и на основе исследований эндосимбионтов было предложено несколько филогенетических отношений.
Eriosomatinae (шерстистая тля)
Aphidina ( Aphis spp, например, бобовая тля )
Анатомия

У большинства тлей мягкое тело, которое может быть зеленым, черным, коричневым, розовым или почти бесцветным. У тлей есть усики с двумя короткими широкими базальными сегментами и до четырех тонких терминальных сегментов. У них есть пара сложных глаз с глазным бугорком позади и над каждым глазом, состоящих из трех линз (называемых триомматидиями). Они питаются соком, используя всасывающий ротовой аппарат , называемый стилетами , заключенный в оболочку, называемую рострумом , которая образована из модификаций нижней и верхней челюсти ротового аппарата насекомых.
У них длинные тонкие лапы с двусачками и двумя лапками . Большинство тлей бескрылые, но у многих видов в определенное время года образуются крылатые формы. У большинства тлей есть пара роговиц (сифункулов) , брюшных трубок на дорсальной поверхности их пятого брюшного сегмента, через которые они выделяют капли быстро затвердевающей защитной жидкости, содержащей триацилглицерины , называемые роговым воском. Некоторые виды также могут продуцировать другие защитные соединения. У тлей над их ректальными отверстиями имеется хвостовидный выступ, называемый хвостом.
Когда качество растения-хозяина становится плохим или условия становятся скученными, некоторые виды тлей производят крылатое потомство (крылатые крылышки ), которое может рассредоточиться по другим источникам пищи. Ротовой аппарат или глаза могут быть маленькими или отсутствовать у некоторых видов и форм.
Рацион питания
Многие виды тлей являются монофагами (то есть питаются только одним видом растений). Другие, такие как зеленая персиковая тля, питаются сотнями видов растений из многих семейств . Около 10% видов в разное время года питаются разными растениями.
Новое растение-хозяин выбирает крылатый взрослый с помощью визуальных сигналов с последующим обонянием с помощью антенн; если растение пахнет правильно, следующим действием будет прощупывание поверхности при посадке. Вставляется игла и выделяется слюна, отбирается проба сока, можно попробовать ксилему и, наконец, исследуют флоэму. Слюна тли может подавлять механизмы запечатывания флоэмы и содержит пектиназы, облегчающие проникновение. Растения, не являющиеся хозяевами, могут быть отклонены на любой стадии исследования, но перенос вирусов происходит на ранней стадии процесса исследования, во время введения слюны, поэтому растения, не являющиеся хозяевами, могут инфицироваться.
Тля обычно питаются пассивно на соке из флоэмы сосудов у растений, как и многие другие hemipterans , такие как щитовки и цикады. После прокола сосуда флоэмы сок, находящийся под давлением, выталкивается в пищевой канал тли. Иногда тля также потребляет ксилемный сок, который представляет собой более разбавленную пищу, чем сок флоэмы, поскольку концентрация сахаров и аминокислот составляет 1% от таковых во флоэме. Ксилемный сок находится под отрицательным гидростатическим давлением и требует активного всасывания, что предполагает важную роль в физиологии тли. Поскольку после периода обезвоживания наблюдается заглатывание ксилемного сока, считается, что тля потребляет ксилемный сок для пополнения своего водного баланса; потребление разбавленного сока ксилемы, позволяющее тле регидратироваться. Однако недавние данные показали, что тля потребляет больше ксилемного сока, чем ожидалось, и это особенно заметно, когда они не обезвожены и когда их плодовитость снижается. Это предполагает, что тля и, возможно, все виды отряда Hemiptera, питающиеся флоэмным соком, потребляют ксилемный сок по причинам, не связанным с пополнением водного баланса. Хотя тля пассивно поглощает сок флоэмы, который находится под давлением, они также могут втягивать жидкость при отрицательном или атмосферном давлении, используя цибариально-глоточный насосный механизм, расположенный в их голове.
Потребление ксилемного сока может быть связано с осморегуляцией . Высокое осмотическое давление в желудке, вызванное высокой концентрацией сахарозы, может привести к переносу воды от гемолимфы в желудок, что приводит к гиперосмотическому стрессу и, в конечном итоге, к гибели насекомого. Тля избегает этой участи, осморегулируя через несколько процессов. Концентрация сахарозы непосредственно снижается за счет ассимиляции сахарозы для метаболизма и за счет синтеза олигосахаридов из нескольких молекул сахарозы , таким образом снижая концентрацию растворенного вещества и, следовательно, осмотическое давление. Затем олигосахариды выводятся через падь, что объясняет его высокую концентрацию сахара, который затем может использоваться другими животными, такими как муравьи. Кроме того, вода переносится из задней кишки , где осмотическое давление уже снижено, в желудок для разбавления содержимого желудка. В конце концов, тля потребляет ксилемный сок, чтобы снизить осмотическое давление в желудке. Все эти процессы действуют синергетически и позволяют тлям питаться соком растений с высокой концентрацией сахарозы, а также адаптироваться к различным концентрациям сахарозы.
Сок растений — несбалансированная диета для тли, поскольку в нем отсутствуют незаменимые аминокислоты , которые тля, как и все животные, не может синтезировать, и обладает высоким осмотическим давлением из-за высокой концентрации сахарозы . Незаменимые аминокислоты доставляются тлю бактериальными эндосимбионтами , обитающими в специальных клетках, бактериоцитах . Эти симбионты перерабатывают глутамат, отходы метаболизма своего хозяина, в незаменимые аминокислоты.
Каротиноиды и фотогетеротрофия
Некоторые виды тлей приобрели способность синтезировать красные каротиноиды путем горизонтального переноса генов от грибов . Это единственные животные, кроме двухпочечного паутинного клеща и восточного шершня, обладающего такой способностью. Используя свои каротиноиды, тля может поглощать солнечную энергию и преобразовывать ее в форму, которую могут использовать их клетки, — АТФ . Это единственный известный пример фотогетеротрофии у животных. В каротине пигменты в тли образуют слой близко к поверхности кутикулы, идеально подходят для поглощения солнечного света. Возбужденные каротиноиды восстанавливают НАД до НАДН, который окисляется в митохондриях для получения энергии.
Размножение
Простейшая репродуктивная стратегия заключается в том, чтобы у тли был один хозяин круглый год. При этом он может чередоваться между половым и бесполым поколениями (голоциклический) или, альтернативно, весь молодняк может быть произведен партеногенезом , яйца никогда не откладываются (анголоциклический). Некоторые виды могут иметь как голоциклические, так и анголоциклические популяции при разных обстоятельствах, но ни один из известных видов тлей не размножается исключительно половым путем. Чередование полового и бесполого поколений могло происходить неоднократно.
Однако размножение тли часто бывает более сложным и включает миграцию между разными растениями-хозяевами. Примерно у 10% видов наблюдается чередование древесных (первичных хозяев), на которых зимует тля, и травянистых (вторичных) растений-хозяев, где они обильно размножаются летом. Некоторые виды могут образовывать касты солдат, другие виды демонстрируют обширный полифенизм в различных условиях окружающей среды, а некоторые могут контролировать соотношение полов в своем потомстве в зависимости от внешних факторов.
Когда используется типичная изощренная репродуктивная стратегия, только самки присутствуют в популяции в начале сезонного цикла (хотя было обнаружено, что у некоторых видов тлей в это время есть и мужской, и женский пол). Из зимующих яиц, которые вылупляются весной, рождаются самки, называемые фундатрисами (стеблевые матери). В размножении обычно не участвуют самцы ( партеногенез ), а в результате рождается живое животное ( живородство ). Живое молодняк производится в результате псевдоплацентарного живорождения, которое представляет собой развитие яиц с дефицитом желтка, зародышей, питающихся тканью, выступающей в роли плаценты. Птенцы появляются из матери вскоре после вылупления.
Яйца партеногенетически производятся без мейоза, а потомство клонально по отношению к своей матери, поэтому все они являются самками ( телитоки ). Эмбрионы развиваются в овариолах матери , которые затем дают рождение живым (уже вылупившимся) нимфам-самкам первого возраста . Поскольку яйца начинают развиваться сразу после овуляции, взрослая самка может содержать развивающиеся нимфы-самки, внутри которых уже есть партеногенетически развивающиеся эмбрионы (т. Е. Они рождаются беременными). Такое телескопирование поколений позволяет тле быстро увеличиваться в численности. Потомство похоже на своего родителя во всем, кроме размера. Таким образом, питание женщины может влиять на размер тела и рождаемость более двух поколений (дочерей и внучок).
Этот процесс повторяется в течение всего лета, производя несколько поколений, которые обычно живут от 20 до 40 дней. Например, некоторые виды капустной тли (например, Brevicoryne brassicae ) могут давать до 41 поколения самок за сезон. Таким образом, одна самка, вылупившаяся весной, теоретически может произвести миллиарды потомков, если все они выживут.

Осенью тля размножается половым путем и откладывает яйца . Факторы окружающей среды, такие как изменение фотопериода и температуры , или, возможно, более низкое количество или качество пищи, заставляют самок партеногенетически производить половых самок и самцов. Самцы генетически идентичны своим матерям, за исключением того, что с системой определения пола X0 тлей у них на одну половую хромосому меньше . У этих половых тлей могут отсутствовать крылья или даже ротовой аппарат. Половые самки и самцы спариваются, а самки откладывают яйца, которые развиваются вне матери. Яйца переживают зиму и следующей весной из них вылупляются крылатые (крылатые) или бескрылые самки. Это происходит в, например, жизненный цикл из розового тли ( Macrosiphum Rosae ), которую можно считать типичной семьи. Однако в теплых условиях, таких как тропики или теплицы , тля может продолжать бесполое размножение в течение многих лет.
Тля, размножающаяся бесполым путем партеногенезом, может иметь генетически идентичное потомство крылатых и не крылатых самок. Контроль сложен; некоторые тли в течение своего жизненного цикла чередуют генетический контроль ( полиморфизм ) и экологический контроль ( полифенизм ) образования крылатых или бескрылых форм. Крылатое потомство, как правило, производится более обильно в неблагоприятных или стрессовых условиях. Некоторые виды производят крылатое потомство в ответ на низкое качество или количество пищи. например, когда растение-хозяин начинает стареть . Крылатые самки мигрируют, чтобы создать новые колонии на новом растении-хозяине. Например, яблочная тля ( Aphis pomi ) после производства многих поколений бескрылых самок дает начало крылатым формам, которые летают на другие ветви или деревья ее типичного пищевого растения. Тля, на которую нападают божьи коровки , златоглазки , паразитоидные осы или другие хищники, может изменить динамику производства своего потомства. Когда эти хищники нападают на тлей, феромоны тревоги, в частности бета-фарнезен , высвобождаются из роговиц . Эти тревожные феромоны вызывают несколько поведенческих модификаций, которые, в зависимости от вида тли, могут включать уход и падение с растения-хозяина. Кроме того, восприятие феромона тревоги может побудить тлей произвести крылатое потомство, которое может покинуть растение-хозяин в поисках более безопасного места для кормления. Вирусные инфекции, которые могут быть чрезвычайно опасными для тлей, также могут привести к появлению крылатого потомства. Например, инфекция Densovirus оказывает негативное влияние на размножение розовой яблонной тли ( Dysaphis plantaginea ), но способствует развитию крылатых тлей, которые могут легче передавать вирус новым растениям-хозяевам. Кроме того, симбиотические бактерии, которые живут внутри тлей, также могут изменять репродуктивные стратегии тлей в зависимости от воздействия факторов окружающей среды.

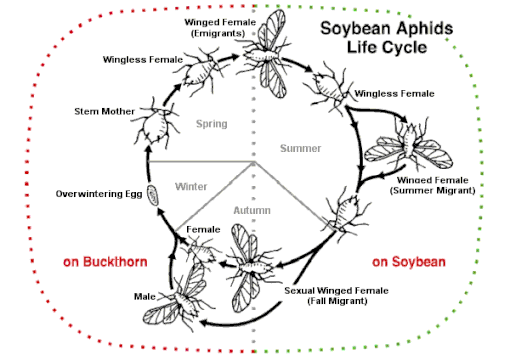
Осенью чередующиеся с хозяевами ( разнородные ) виды тлей образуют особое крылатое поколение, которое летает на разные растения-хозяева в течение половой части жизненного цикла. Производятся нелетающие женские и мужские половые формы, откладывающие яйца. Некоторые виды, такие как Aphis fabae (бобовая тля), Metopolophium dirhodum (розовая тля), Myzus persicae (персиково-картофельная тля) и Rhopalosiphum padi (черемуха-овсяная тля), являются серьезными вредителями. Зимуют на основных хозяевах на деревьях или кустах; летом они мигрируют к своему вторичному хозяину на травянистом растении, часто урожае, затем осенью гинопары возвращаются на дерево. Другой пример — соевая тля ( Aphis glycines ). По мере приближения осени растения сои начинают стареть снизу вверх. Тля выталкивается вверх и начинает давать крылатые формы, сначала самок, а затем самцов, которые улетают к основному хозяину — крушине . Здесь они спариваются и зимуют как яйца.
Источник