
Тля клещи однолетние травянистые растения организмы с экспоненциальной кривой роста




12.6.3. Рост популяции и кривые роста
Если рождаемость в популяции превышает смертность, то популяция будет расти, если, конечно, изменения в результате иммиграции и эмиграции незначительны. Чтобы понять закономерности роста популяций, полезно вначале рассмотреть модель (см. гл. 2), описывающую рост популяции бактерий после посева их на свежую культуральную среду. В этой новой и благоприятной среде условия для роста популяции оптимальны и наблюдается экспоненциальный рост (см. рис. 2.7; см. также разд. 21.1.2). Кривая такого роста — это экспоненциальная, или логарифмическая кривая. Но, как сказано в разд. 2.2.4 и 21.1.2, в конце концов достигается такая точка, когда по нескольким причинам, в том числе из-за уменьшения пищевых ресурсов и накопления токсичных отходов метаболизма экспоненциальный рост становится невозможным. Он начинает замедляться так, что кривая роста приобретает сигмоидную (S-образную) форму, как показано на рис. 2.7 для бактерий и на рис. 21.1 для дрожжей. Такой тип роста называют зависимым от плотности, так как скорость роста зависит от плотности популяции, которая влияет на истощений пищевых ресурсов и накопление токсичных продуктов, а потому на рост. С увеличением плотности скорость роста популяции постепенно снижается до нуля, а кривая выходит на плато. При нулевом росте популяция стабильна, т. е. размеры ее не меняются (напомним, что отдельные организмы при этом могут расти и размножаться; нулевая скорость роста популяции означает лишь то, что скорость размножения, если оно происходит, уравновешена смертностью). Такая сигмоидная кривая роста получена для ряда одноклеточных и многоклеточных организмов, например для клеток водорослей в культуральной среде, для фитопланктона озер и океанов весной, для насекомых, таких, как мучные хрущаки или клещи, интродуцированные в новое местообитание с обильными запасами пищи, где нет хищников.
Кривая другого типа получается, когда экспоненциальный рост продолжается вплоть до внезапного падения плотности популяции в результате исчерпания ресурсов среды. Эту кривую называют «J-образной» или кривой типа «бум и крах». Такой рост не зависит от плотности, так как его регуляция не связана с плотностью популяции до самого момента катастрофы. Крах может происходить по тем же причинам, например из-за истощения пищевых ресурсов, которое в случае сигмоидной кривой роста заблаговременно оказывало регулирующее влияние на рост. Миграция или расселение, так же как и внезапное снижение скорости размножения, может способствовать уменьшению численности популяции. Расселение может быть связано с определенной стадией жизненного цикла, например с образованием семян. На рис. 12.33 показаны примеры того и другого типа. Для обоих типов характерна экспоненциальная фаза в начале роста.
Рассматривая вопрос об оптимальных размерах популяции в данной среде, важно учитывать поддерживающую емкость, или «кормовую продуктивность», этой среды. Чем выше поддерживающая емкость, тем больше максимальный размер популяции, который может существовать в данном местообитании неопределенно долгое время. Дальнейшему росту популяции будет препятствовать один или несколько лимитирующих факторов. Это зависит от доступности ресурсов для данного вида. Как показано на рис. 12.33, в случае J-образной кривой роста популяция внезапно выходит за пределы поддерживающей емкости среды. Эту величину обозначают символом К, который можно использовать также для обозначения максимальных размеров стабильной популяции в данных условиях. Рост, соответствующий сигмоидной и J-образной кривым, можно описать алгебраически с помощью простых дифференциальных уравнений (оба уравнения относятся к популяциям, в которых поколения полностью перекрываются, так что популяция изменяется непрерывно; это и позволяет использовать дифференциальные уравнения). Такие уравнения приведены в табл. 12.10, там же дается краткое объяснение.
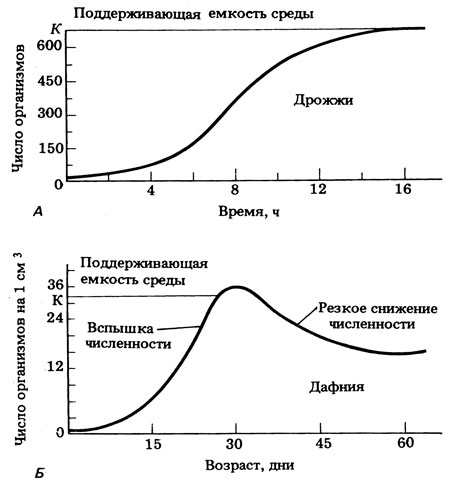
Рис. 12.33. Два типа кривых роста популяции. (А) Сигмоидная (S-образная) кривая роста дрожжей в культуре. Поддерживающая емкость среды определяется изначальным количеством доступных питательных веществ, содержащихся в среде. (Б) J — образная кривая роста дафний в культуре (A.S. Boughey (1971). Fundamental Ecology, International Textbook Co.) См. также рис. 12.40-пример роста популяции оленей на плато Кайбаб
| Если N — число особей в популяции и t — время, то скорость изменения численности во времени, представлена отношением dN/dt, пропорциональна N, т. е. dN/dt =rN, где r константа — врожденная скорость роста численности популяции, связанная с максимальной скоростью размножения особи данного вида. Чем выше скорость размножения, тем больше значение r | |||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| J — образная кривая (рост не зависящий от плотности) | Сигмоидная кривая (рост, зависящий от плотности) | ||||||||||||||||||||||||||||||||||||||||||
| r — Виды (виды — «оппортунисты») | K — виды (с тенденцией к равновесию) |
|---|---|
| Размножаются быстро (высокая плодовитость, время генерации короткое), поэтому значение r (врожденная скорость роста популяции) высокое | Размножаются медленно (низкая плодовитость, продолжительное время генерации), поэтому значение r низкое |
| Скорость размножения не зависит от плотности популяции | Скорость размножения зависит от плотности популяции, быстро увеличивается, если плотность падает |
| Энергия и вещество распределяются между многими потомками | Энергия и вещество концентрируются в немногих потомках; родители заботятся о потомстве |
| Размеры популяции некоторое время могут превышать К (поддерживающую емкость среды) | Размеры популяции близки к равновесному уровню, определяемому К |
| Вид не всегда устойчив на данной территории | Вид устойчив на данной территории |
| Расселяются широко и в больших количествах; у животных может мигрировать каждое поколение | Расселяются медленно |
| Размножение идет с относительно большими затратами энергии и вещества | Размножение идет с относительно малыми затратами энергии и вещества; большая часть энергии и вещества расходуется на нерепродуктивный (вегетативный) рост |
| Малые размеры особей | Крупные размеры особей; у растений деревянистые стебли и большие корни |
| Малая продолжительность жизни особи | Большая продолжительность жизни особи |
| Могут поселяться на открытом грунте | Плохо приспособлены к росту на открытых местах |
| Местообитания сохраняются недолго (например, зрелые фрукты для личинок Drosophila) | Местообитания устойчивые и сохраняются долго (например, лес для обезьян) |
| Слабые конкуренты (способность к конкуренции не требуется) | Сильные конкуренты |
| Защитные приспособления развиты сравнительно слабо | Хорошие защитные механизмы |
| Не становятся доминантами | Могут становиться доминантами |
| Лучше приспособлены к изменениям окружающей среды (менее специализированные) | Менее устойчивы к изменениям условий среды (высокая специализация для жизни в устойчивых местообитаниях) |
| Примеры | Примеры |
| Бактерии | Крупные тропические бабочки |
| Парамеция | Кондор (крупная хищная птица) |
| Тли | Альбатрос |
| Мучные хрущаки | Человек |
| Однолетние растения | Деревья |
По существу, эти две стратегии представляют два различных решения одной задачи — задачи длительного выживания вида. Виды с r-стратегией быстрее заселяют нарушенные местообитания (характерные для ранних стадий сукцессий, такие, как обнаженная горная порода, лесные вырубки, выгоревшие участки), чем виды с Х-стратегией, так как они легче распространяются и быстрее размножаются. Виды с Х-стратегией более конкурентоспособны, и в конце концов они вытесняют r-виды, которые тем временем перемещаются в другие нарушенные местообитания. Высокий репродуктивный потенциал r-видов означает, что, оставшись в каком-либо местообитании, они быстро использовали бы доступные ресурсы и превысили поддерживающую емкость среды, а затем популяция погибла бы. Другими словами, для них характерна J-образная кривая роста с быстрым падением численности популяции в конце (рис. 12.33, Б). Виды с r-стратегией занимают данное местообитание лишь в течение жизни одного или, самое большее, нескольких поколений. Затем они переселяются на новое место. Отдельные популяции могут регулярно вымирать, но вид при этом перемещается и выживает. В целом эту стратегию можно охарактеризовать как стратегию «борьбы или бегства».
Как отмечает Саутвуд * , хотя для самых мелких организмов, таких, как бактерии и насекомые, характерна r-стратегия, а для самых крупных, таких, как многие позвоночные и деревья,- Х-стратегия, большинству организмов свойственны стратегии промежуточных типов. Даже в пределах таких групп, как насекомые и позвоночные, можно обнаружить различные стратегии. Среди птиц, например, лазоревка (оппортунистический обитатель лиственных лесов) проявляет резко выраженную r-стратегию. Будучи маленькой, она тем не менее откладывает сравнительно много яиц и способна более чем вдвое увеличить свою численность за один сезон. У одного из самых мелких попугаев — волнистого попугайчика, кочующего по Центральной Австралии,- очень небольшое время генерации, поэтому этот вид тоже можно отнести к видам с r-стратегией. Но существуют и птицы с резко выраженной К-стратегией, например кондор и альбатрос. Размах крыльев у них превышает три метра, но они откладывают только по одному яйцу каждые два года. Альбатрос позднее всех других птиц достигает половой зрелости (к 9-11 годам).
* ( Т. R. Е. Southwood, Bionomic Strategies and Population Parameters, in: Theoretical Ecology. Principles and Applications, ed. R.M. May (1976), Blackwell.)
Стабильные местообитания способствуют отбору на К-стратегию. Хорошим примером служат мясные (падальные) мухи, личинки которых питаются падалью. У одного вида (Blaseoxiphia fletcheri) ли-чинки живут в жидкости кувшинчатых листьев насекомоядного растения саррацении — более долговечном местообитании, чем падаль, которую используют другие виды. Самка этой мухи производит всего лишь 11 личинок (тогда как самки родственных видов — от 50 до 170); каждая такая личинка имеет сравнительно крупные размеры.
12.19. Изучите рис. 12.34, где представлена доля цветков в общей биомассе на корню у трех видов золотарника (Solidago) на востоке США. На рисунке указано, где растет каждый из этих видов — в лесу или на открытых сухих местах (в нарушенных местообитаниях на ранних стадиях сукцессии):
а. Какие виды расходуют на размножение больше биомассы?
б. Какие виды в большей степени подвержены г-отбору?
в. Какие виды в большей степени подвержены К-отбору?
г. Какой общий вывод можно сделать, сравнив интенсивность цветения S. speciosa в лесу и на открытых сухих участках?
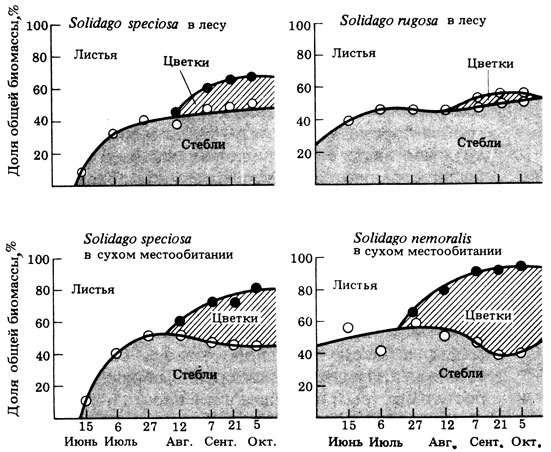
Рис. 12.34. Процент биомассы, приходящийся на листья, стебли и цветки в четырех популяциях золотарника (R. М. May (ed.) (1976). Teoretical Ecology, Blackwell.)
Обращайтесь к шлюхам в любое время дня и приглашайте их к себе в дом. Только хорошие проститутки из вашего района с счастьем готовы испытать удовольствие и заняться благополучным трахом.
Источник