
Чередование поколений
Имеется несколько групп насекомых, последовательные поколения у которых существенно отличаются друг от друга по способу размножения, а иногда и по поведению.
Насекомые, у которых репродуктивной функцией наделены только взрослые особи. К этой группе относятся растительноядные насекомые: тли и галло-образующие осы.
Тли (Aphididae, Hemiptera) характеризуются сложными и варьирующими сезонными циклами со сменой поколений, размножающихся половым путём с откладкой яиц и партеногенетических с живорождением, крылатых и бескрылых, а также миграциями с летнего растения-хозяина на зимнее.
Наиболее простой сезонный цикл у капустной тли Brevicoryne brassicae. Зимующей фазой у этих тлей являются яйца, отложенные осенью на стебли крестоцветных растений. Личинки, вылупившиеся из этих яиц следующей весной, развиваются в бескрылых партеногенетических живородящих самок — тлей-основательниц. Интересно отметить, что все яйца дают самок-основательниц. От этих особей получается поколение партеногенетических живородящих самок, которые могут быть как крылатыми, так и бескрылыми. Такие поколения живородящих самок появляются одно за другим в течение лета. Обычно партеногенетические особи живут около месяца и рождают от 50 до 100 личинок. Осенью, когда дни становятся короче, живородящие самки дают поколение, состоящее из бескрылых самок и крылатых самцов, размножающееся половым путём. После спаривания каждая самка откладывает от одного до нескольких яиц, которые остаются на зиму.
Более сложные сезонные циклы включают ещё ряд особенностей. Многие виды мигрируют на летние растения-хозяева. У таких тлей зимнее, или первичное, растение-хозяин обычно много-летнее. Яйца откладываются на это растение осенью, личинки выходят из яиц весной, и из них развиваются бескрылые тли-основательницы. Эти самки дают начало поколению бескрылых живородящих самок, от которых получается поколение крылатых живородящих самок, перелетающих на летнее, или вторичное, растение-хозяина. Поколения живородящих самок, крылатых или бескрылых, появляются на этом растении вплоть до осени, когда рождается поколение мигрирующих особей, перелетающих на зимнее растение; здесь появляется молодое поколение, состоящее из самок и самцов.
У некоторых видов тлей крылатые самцы рождаются на летнем растении, мигрируют на зимнее, где спариваются с бескрылыми яйцекладущими самками. У других видов и самки, и самцы — бескрылые и происходят от крылатых живородящих самок, мигрировавших ранее на зимнее растение-хозяина. У некоторых видов тлей разные поколения резко отличаются друг от друга по своему поведению. Например, в роде Pemphigus самка-основательница делает галл на листовой пластинке или черешке листа тополя. Её потомки мигрируют на корни сложноцветных и других растений и дают начало нескольким поколениям, живущим на корнях и не образующим галлов.
Филлоксеры (Phylloxeridae, Hemiptera) близки к тлям; у них также наблюдается чередование партеногенетических и половых поколений (несколько крылатых и несколько бескрылых) и сложные миграции. От тлей они, однако. отличаются тем, что все самки яйцекладущие. Наиболее типичным примером является виноградная филлоксера (рис. 7.19). Ряд поколений партеногенетических самок образуют галлы на листьях растений, а осенью мигрируют на корни винограда, где следующей вес- ной новые партеногенетические поколения вызывают образование галлов на мелких корнях винограда. Поскольку насекомые в этих поколениях происходят от неоплодотворённых особей, они называются агамными. Некоторые особи, живущие на корнях, дают осенью начало партеногенетическим агамным крылатым мигрантам, выползающим из земли и перелетающим на виноградную лозу. Появившееся здесь потомство состоит из бескрылых агамных самцов и самок. После спаривания каждая самка откладывает одно яйцо в трещину коры. Яйца перезимовывают, следующей весной из них вылупляются личинки, дающие начало серии поколений, живущих на листьях.
Среди галлообразующих ос (Cynipidae, Hymenoptera) имеется много видов, которым свойственно чередование половых и партеногенетических поколений. У Andricus erinacei, живущей на дубе, зимуют яйца, отложенные в листовые или цветочные почки. Вылупление из яиц происходит весной, и каждая личинка оказывается окружённой мягким почечным галлом. Внутренний слой галла личинка использует в качестве пищи. Ранним летом по завершении личиночного развития появляются крылатые самцы и самки. Самки откладывают яйца на жилки листьев дуба. Вылупившиеся из яиц личинки вызывают аномальное разрастание жилок листа с образованием галлов, которые носят название галлов-ёжиков. Личинки в этих галлах достигают зрелости осенью; из них развиваются самки с короткими крыльями, которые откладывают партеногенетические зимующие яйца в дубовые почки.
Чередование поколений, среди которых есть партеногенетические, создаёт возможность для быстрого роста численности популяции, что обеспечивает успешное использование короткого периода благоприятных условий. Благодаря партеногенезу отдельные самки могут давать начало новым популяциям, быстро расселяясь с помощью ветра, или, наоборот, снижать численность при неблагоприятных условиях.
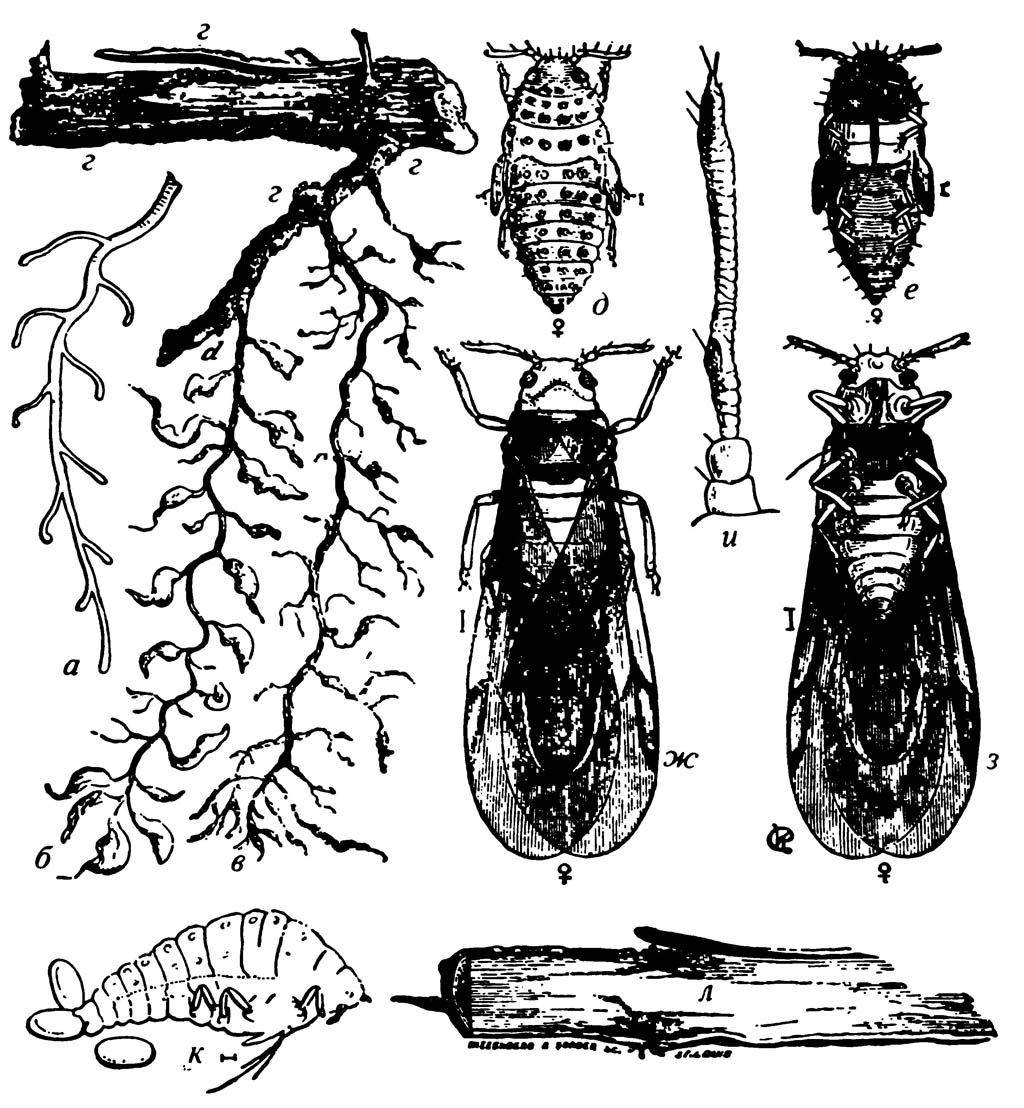
7.19. Виноградная филлоксера; форма, живущая на корнях винограда
а — здоровый корень; б — корень, поражённый насекомыми; видны узелки и вздутия, вызванные их уколами; в — корень, покинутый насекомыми, с начинающими отмирать корешками; г — насекомые на крупном корне; д — агамная женская нимфа, вид с дорсальной стороны; е — та же нимфа с вентральной стороны; ж — крылатая агамная самка, вид с дорсальной стороны; з — та же самка с вентральной стороны; и — увеличенная антенна крылатого насекомого; к — бескрылая агамная самка, кладущая яйца на корни; л — гниение большого корня в результате проколов насекомых
Источник
Партеногенез

Как известно, в основе полового размножения лежит процесс оплодотворения. Мужские клетки сливаются с женскими, в результате чего образуется зародыш, из которого развивается организм, несущий в себе признаки отца и матери. Партеногенез — удивительный вариант полового размножения, когда новая жизнь возникает без малейшего участия самца. Потомство появляется из неоплодотворенных половых клеток самки и, по сути, становится ее копией. Это феномен в животном царстве представлен довольно широко даже среди позвоночных: например, подобным образом размножаются некоторые ящерицы. Однако же отсутствие оплодотворения чаще встречается у тех организмов, которые находятся внизу эволюционного древа, то есть у беспозвоночных. Среди всех групп насекомых к партеногенезу более всего склонны равнокрылые.
Кто такие равнокрылые?
Традиционно в отряд равнокрылых относят цикад, а также опасных вредителей растений — тлей, кокцид и листоблошек. Партеногенез характерен только для представителей трех последних групп, но в целом всех их объединяет общее строение крыльев, в частности их твердость. Тем не менее в последнее время ученые засомневались, стоит ли выделять данных насекомых в особый отряд, возможно, их следует отнести к полужесткокрылым (клопам), с которыми у них много общего.
Жизненный цикл тлей
Принцип партеногенетического размножения лучше всего рассматривать на примере тлей. Данные насекомые имеют очень сложный жизненный цикл, который начинается с зимующих яиц. Весной из этих яиц на свет выходят лишенные крыльев самки и размножаются сами по себе (то есть партеногенезом). Так путем живорождения появляется несколько поколений аналогичных бескрылых насекомых. Однако в определенное время в очередном выводке самки рождаются крылатыми — у одних видов они перелетают на другое растение, у других остаются на прежнем месте. Потом вновь идет безудержное размножение женских особей с помощью все того же партеногенеза. Иногда у отдельных видов за лето появляется до 17 поколений, причем некоторые особи крылаты и способны к миграциям. С наступлением осени впервые за год рождаются крылатые самцы. Теперь возможно настоящее половое размножение с участием обоих полов, после которого самки сумеют отложить яйца на зиму.

Партеногенез: достоинства и недостатки
Партеногенез сводит на нет главное преимущество полового размножения — повышение генетического разнообразия потомков, которое достигается за счет того, что каждый организм получает ДНК обоих родителей. В данном же случае происходит почти полное копирование наследственного материала матери, а такая неизменность генофонда должна в конце концов привести к вырождению популяции. Однако в строгой форме (когда самки в течение многих поколений воспроизводят исключительно себе подобных) партеногенез встречается лишь у более примитивных животных. И даже в этом случае природа разработала сложные и до сих пор не совсем понятные механизмы обогащения новыми генами.
.jpg)
У насекомых партеногенез является лишь этапом жизненного цикла, который со временем и сменяется нормальным двуполым размножением. В таком случае его преимущества становятся очевидны. Во-первых, гораздо быстрее растет численность, ведь для размножения не нужны самцы, а значит, насекомые не ищут себе пары и не выполняют сложных брачных ритуалов, сражаясь за партнера. Во-вторых, для появления новой популяции достаточно всего одной-единственной материнской особи.
Источник
Энциклопедия «Жизнь животных» (1970)
ПОДОТРЯД ТЛИ (APHIDODEA)
ПОДОТРЯД ТЛИ (APHIDODEA)
Тли, бесспорно, очень интересная группа насекомых и по своей исключительно разнообразной биологии, и по обилию видов, и по тому значению, которое они имеют как вредители. Это мелкие нежные насекомые, обычно сиДящие большими колониями либо на надземных частях растений, либо на корнях. Многие тли вызывают болезненные изменения тех частей растений, на которых сидят, вызывая скручивание листьев, деформацию концов побегов, а иногда и образование полых внутри выростов — галлов. Многие тли выделяют сладкие экскременты, называемые медвяной росой, благодаря чему их охотно посещают муравьи, питающиеся этими выделениями. Не все тли выделяют медвяную росу. Ее не выделяют виды, тело которых покрыто восковым пушком, и обитатели галлов.
Причина выделения медвяной росы состоит в том, что покровы у тлей нежные и через них легко испаряется вода. Поэтому, чтобы не погибнуть от высыхания, тли много «пьют», поглощают много соков растений, на которых питаются. А питье тли — это и пища; сока тля поглощает больше, чем ей нужно для питания, избыток неусвоенных растворенных в соке растений Сахаров выводится в непереваренном виде. В жарких сухих районах, например в пустынях, тли выделяют гораздо больше медвяной росы, чем во влажных местностях. Поэтому понятно, что защищенные от высыхания восковым налетом или живущие внутри замкнутого галла тли не выделяют сахара — они меньше «пьют» сока, потребляя его лишь столько, сколько надо для питания.
У тлей очень сложный цикл развития. Наиболее типичный цикл развития с зимующими оплодотворенными яйцами. Весной из яиц выходят бескрылые самки, которые размножаются без оплодотворения (партеногенетически). Партеногенетически размножающиеся самки у большинства тлей яиц не откладывают, а рождают живых детенышей. Правда, это не настоящее живорождение. При настоящем живорождении (у млекопитающих, а также у скорпионов, перипатов) зародыш развивается за счет веществ, постепенно поступающих из организма матери. А у тлей наблюдается яйцеживорождение: партеногенетическая тля яиц не откладывает, а маленькая тля развивается из яйца за счет желтка, но только развитие протекает в организме матери.
Бывают тли, весь цикл развития которых протекает на одном виде растений (одноядные тли), бывают и такие, которые развиваются на разных видах растений (многоядные тли). Среди многоядных есть такие, для которых смена видов растений не обязательна, но у многих видов тлей происходит обязательная смена кормовых растений — это мигрирующие тли. В последнем случае тот вид растения, на котором перезимовывают яйца, называется основным кормовым растением. Вышедшие из яиц партеногенетические бескрылые самки называются основательницами. Иногда несколько бескрылых поколений потомства основательницы остается на том же растении, и лишь впоследствии среди них появляются крылатые тли (тоже партеногенетические самки), а иногда уже и первое поколение состоит из партеногенетических самок, перелетающих на другие растения. Это так называемые расселительницы. Переселяясь на новые кормовые растения, самки-эмигранты рождают бескрылых потомков, образующих новые колонии. Затем в потомстве бескрылых появляются снова крылатые особи, способные к дальнейшему расселению. Поколения тлей, развивающихся на дополнительных видах кормовых растений, т. е. на тех, на которые перекочевали расселительницы, называются переселенцами.
За лето многие тли дают несколько поколений. К осени среди переселенцев появляются так называемые полоноски, которые рождают самцов и самок. Оплодотворенные самки перелетают на те виды растений, на которых весной бывают основательницы, и откладывают зимующие, покрытые плотной оболочкой яйца.
Такой тип смены поколений у тлей очень обычен, но бывают и другие циклы. Разбор конкретных примеров может это хорошо иллюстрировать. Так как у тлей развивается несколько поколений в году, они размножаются с поразительной быстротой. Подсчитано, что в тех субтропиках, где около 300 теплых дней в году, потомство одной тли вырастает в 10/15 раз! Конечно, такого обилия потомков никогда не бывает. Но практика борьбы с вредителями, например, садовых культур показывает, что, если не вести борьбу, тли могут почти сплошь покрывать растения, а от многих тлей происходит гибель возделываемых растений.

Одни из самых примитивных тлей — хермесы. У желтого хермеса (Chermes abietis), развивающегося на ели, крылатые особи появляются во второй половине лета и откладывают яйца на хвое у основания почек. Из яиц выходят зимующие основательницы, вызывающие образование галлов (рис. 206, 4). Этот вид, немигрирующий, партеногенетический, дает одно поколение в году. А очень близкий вид — зеленый хермес (Ch. viridis) — тоже перезимовывает в виде самок-основательниц на ели, но у него крылатые особи перелетают на лиственницу, где и откладывают яйца. Из них выходят тли, которые на коре лиственницы зимуют, а на следующий год кладут тут же яйца. Из яиц выходят крылатые тли-полоноски, а из яиц полоносок выходят самцы и самки. Оплодотворенные самки перелетают обратно на ель и кладут там яйца, из них выходят зимующие основательницы.

К хермесам близка виноградная филлоксера (Phylloxera vastatrix) — тля американского происхождения, которая у себя на родине вредного значения не имела, а попав в Европу, изменила свою биологию и стала опаснейшим вредителем, настоящим бичом виноградарства. Сперва она была завезена во Францию и разорила виноградное хозяйство этой страны, славящейся своим виноделием. Распространилась она и далее на восток. За 30 лет она погубила около 6 миллионов гектаров виноградников! У нас филлоксера есть не во всех районах возделывания винограда — только в Молдавии, на Правобережье УССР, на Северном Кавказе и местами в Закавказье.
Огромный вред, причиненный в Европе филлоксерой, объясняется тем, что европейская лоза, ранее не встречавшаяся с этим вредителем, сильно от него страдает, тогда как американская, эволюция которой шла бок о бок с эволюцией этого вредителя, вырабатывает защитные реакции против филлоксеры. Казалось бы, что наиболее простой путь в виноградарстве — замена европейского винограда американским. Но дело в том, что у американской лозы все показатели (урожайность, сахаристость, вкусовые качества ягод) намного ниже, чем у давно культивируемых сортов европейской лозы.
В чем же особенности развития американской филлоксеры на американской лозе? В Америке на американской лозе из перезимовавшего на коре яйца выходят основательницы — мелкие (0, 7 мм) желтые тли, переползающие на молодые листья винограда. Вокруг сосущей тли образуются маленькие (с гречишное зерно) галлы. Основательница бескрылая. Она кладет много яиц, из них выходят снова личинки, тоже вызывающие на листьях галлы. Это поколение листовых филлоксер снова кладет яйца, из которых частично выходят такие же личинки, как и те, из которых развились их родительницы, а из последних порций яиц выходят личинки с несколько более толстыми усиками. Эти личинки переползают на корни и, высасывая из них соки, вызывают их болезненные разрастания. Несколько партеногенетических поколений тлей развивается на корнях, где они и зимуют иногда на большой глубине (1 м).
Весной перезимовавшие самки снова кладут на корнях яйца, и так развивается несколько партеногенетических поколений. В жаркое время года, среди лета, часть личинок корневой формы филлоксеры дает оранжевых «нимф» с зачатками крыльев, выходящих из почвы и превращающихся в крылатых самок. Это самкиполоноски. Они откладывают яйца двух типов — мелкие (из них выходят самцы) и крупные (из них выходят самки). И самцы и самки бескрылы и имеют неразвитый ротовой аппарат. После оплодотворения самка откладывает на кору лозы одно яйцо, которое и зимует. Вышедшая из яйца личинка переходит на листья, и цикл повторяется (рис. 208). Американской виноградной лозе корневая форма филлоксеры не приносит большого вреда; в месте сосания довольно быстро образуется защитная пробковая ткань, ограничивающая и возможности питания корневых форм филлоксеры, и их размножение. А питание на листьях большого вреда растению не причиняет. Этим объясняется то, что американским лозам филлоксера не страшна.

А на европейской лозе картина иная; на ней филлоксера не может вызывать образование галлов, если галлы и появляются, то листовое поколение быстро погибает. Но зато на корнях корневая форма развивается великолепно: сосание этих тлей приводит к болезненным изменениям корней, вызывающим усыхание всего растения. Защитные реакции европейских форм винограда против нового для них врага — филлоксеры незначительны. Поэтому на европейской лозе размножение корневой филлоксеры происходит быстро. Хотя корневая форма и дает время от времени на европейской лозе половые особи, они не имеют значения, их потомство погибает, так как листья европейских сортов негодны для развития филлоксеры. Поэтому половое поколение — «рудимент» в жизненном цикле филлоксеры на европейской лозе, где все размножение вредителя осуществляется партеногенетически.
Интересно, что за короткий срок (десятки лет для эволюции короткий срок!) в Европе изменились наследственные свойства филлоксеры. С европейской лозы филлоксера не переходит обратно на американскую — известный «закон необратимости эволюции» бельгийского ученого-палеонтолога Д о л л о оказывается справедлив и для такого короткого срока.
Большой урон, который терпело виноградарство от филлоксеры, заставлял и заставляет и виноградарей, и энтомологов искать решения этой проблемы. Правда, дешевых и радикальных мер борьбы с филлоксерой еще нет — премия, объявленная за их разработку Французской академией наук, еще никому не присуждена! Но разработано много мероприятий, позволивших восстановить виноградное хозяйство Франции и разводить виноград в других местах, где есть филлоксера, правда, с несколько худшими результатами, чем на землях, свободных от нее. Там широко применяют привитую культуру. На устойчивые к корневой филлоксере черенки американской лозы прививают европейские сорта, обеспечивающие высокое качество урожая. Это мероприятие плохо тем, что привитая культура несколько менее урожайна, чем корнесобственная, а кроме того, при этом методе очень удорожается разведение винограда: прививка — трудоемкий процесс.
Рекомендуют сажать виноград на песках — песчаных почв филлоксера не выносит. Найдены некоторые более устойчивые сорта европейских виноградных лоз (ркацители, papa нягру и др.), дольше сопротивляющиеся филлоксере. Предлагаются методы подтравливания филлоксеры на корнях лозы теми ядами, которые в применяемых дозировках не губят лозу (кубовые остатки дихлорэтана и др.)Многого можно ожидать от селекционных работ, сопровождаемых тонкими биохимическими и анатомическими исследованиями растений, организованных в Молдавии академиком Я. И. Принцем.
Но очень важно помнить, что, так как расселяется только корневая филлоксера — нежное, нестойкое и малоподвижное насекомое, важнейшая задача — не завезти филлоксеру в те районы, где ее нет. А таких районов у наемного: Крым, Средняя Азия, часть Закавказья. Внутренний карантин растений в борьбе с филлоксерой имеет огромное значение.
Среди других тлей американского происхождения большой вред нашему садоводству причиняет кровяная тля (Eriosoma lanigerum). Эта тля выделяет густой шерстевидный покров из восковых нитей, скрывающий ее красноватое тело. Если такую тлю раздавить, остается красное пятно, отсюда и название этой тли.

В Америке жизнь этой тли протекает так: на вязе в трещинах коры зимуют яйца. Весной личинки поднимаются к почкам вяза и высасывают молодые листочки, которые при этом скручиваются, слепляясь в розетки. Так развивается 2 поколения бескрылых партеногенетических тлей. Затем появляются крылатые переселенцы («эмигранты»), перелетающие на яблоню. Здесь они рождают личинок, поселяющихся на концах побегов и на листьях. Из этих личинок вырастают бескрылые партеногенетические самки. Их потомки (тоже партеногенетические самки) спускаются вниз на корни, вызывая на них вздутия и галлы. При массовом размножении кровяной тли на корнях яблони засыхают. При этом часть тлей остается на листьях. Они рождают крылатых полоносок, которые перелетают назад на американский вяз и там рождают самцов и самок. Оплодотворенные самки на коре вяза опять откладывают зимующие яйца. Таков полный цикл развития кровяной тли. Но она может непрерывно размножаться и только на яблоне. Именно так и размножается кровяная тля в Европе, куда она была завезена с посадочным материалом и где нет тех видов вязов, на которых она могла бы развиваться. Интересно, что примерно за 200 лет существования в Европе, в местах, где теперь высажен американский вяз, эта тля утратила способность давать полоносок. Поэтому «европейская» кровяная тля не мигрирует.
Кровяная тля — очень опасный вредитель. Ее большие колонии, губящие молодые листья, как густыми хлопьями белой ваты покрывают побеги. На корнях образуются желваки, которые, растрескиваясь, дают широкий доступ возбудителям гнилей. Химическая борьба с кровяной тлей и затруднительна (тля либо в почве, либо под защитой несмачиваемого воскового пушка), и неэффективна. Наилучшие результаты дал завоз ее естественного врага — наездника афелинуса (Aphelinus mali), откладывающего яйца в кровяную тлю и сдерживающего размножение этого грозного вредителя семечковых плодовых культур (рис. 210).

Кровяная тля — теплолюбивое насекомое и встречается у нас в Крыму, в Закавказье и в Молдавии.
Примером нашей немигрирующей тли может служить зеленая яблоневая тля (Aphis pomi), которую можно встретить во всех местностях, где растут яблоня, боярышник, айва или груша. Это бледнозеленая тля с двумя хвостовыми, так называемыми соковыми, трубочками, находящимися на спинной стороне близ вершины брюшка. Осенью оплодотворенные самки откладывают яйца на молодые побеги у основания почек. Когда весной почки набухают, вышедшие из яиц личинки переходят на них, а после распускания почек забираются между листочками. В течение всего лета тля размножается партеногенетически, давая от 7 (в северных районах) до 17 (на юге) поколений. Иногда колонии тли бывают очень многочисленными, насекомые сплошь осыпают листья. Осенью в потомстве тли появляются самцы и самки, после спаривания откладывающие зимующие яйца. Яблоневую тлю охотно посещают муравьи, слизывающие сладкие, как у большинства открытоживущих и незащищенных восковым налетом тлей, экскременты.

Бороться с зеленой яблоневой тлей несложно: своевременно проводимые мероприятия, такие, как весеннее опрыскивание (до раскрытия почек!) эмульсией карболинеума, дают прекрасные результаты. Активно уничтожают яблоневую тлю и многие насекомые-хищники — божьи коровки и их личинки (рис. 211), личинки златоглазок и др., чем сдерживают размножение этого вредителя.
Примером нашей мигрирующей тли может служить черная свекловичная тля (Aphis fabae). Из яиц, зимующих на коре ветвей бересклета возле листовых почек, выходят бескрылые основательницы, которые начинают быстро размножаться на листьях, давая на бересклете несколько поколений. Когда листья бересклета начинают загрубевать, появляются крылатые эмигранты, которые перелетают на свеклу или конские бобы. На листьях этих растений свекловичная тля дает еще 10-12 поколений не только бескрылых, но и крылатых особей, которые расселяются по полям. Поврежденные листья скручиваются, желтеют и сохнут. Осенью появляются самцы и самки, которые откладывают оплодотворенные яйца снова на бересклете.

Есть среди наших тлей и такие, которые с основного кормового древесного растения мигрируют на корни разных травянистых. Так, например, мешковидные стручкообразные галлы на листьях береста вызывают основательницы берестово-злаковой тли (Tetraneura ulmi). Бескрылые переселенцы этой тли развиваются на корнях злаков. На корнях злаков партеногенетические переселенцы могут зимовать, и потому в местностях, где нет береста, может развиваться только так называемая неполноциклая форма этой тли — злаковая корневая тля.
Живущие на корнях переселенцы представляют очень интересные объекты исследований. Примером может служить белая корневая тля (Trifidaphis phaseoli, рис. 213). Летом в Средней Азии на посевах хлопчатника можно встретить завядающие растения. Выкопав такое растение, с первого взгляда на корнях его нельзя обнаружить никаких повреждений. Но если вырыть растение вместе с комом земли и разломать этот ком по ходу корня, на тонких питающих боковых корешках видны нежные мелкие (около 1, 3 мм) белые почти шаровидные насекомые — тли.

Тело белой корневой тли покрыто мелкой восковой пыльцой. Усики у нее короткие, с коротким тупым концом последнего членика. Белая корневая тля принадлежит к числу двудомных мигрирующих тлей: ее основным кормовым растением, на котором зимуют яйца, отложенные осенью оплодотворенными самками, является в предгорных районах Средней Азии фисташка. Вышедшие весной из яиц личинки тли внедряются в листовую почку и начинают сосать развивающийся лист. Под влиянием слюны тли, содержащей ростовые вещества, пластинка листа неправильно разрастается, на краю листа образуется веретеновидный галл, под защитой которого развивается тля. Когда тля заканчивает развитие, галл лопается и крылатая тля-эмигрант перелетает на корни различных двудольных растений, как дикорастущих, так и культурных.
Дальнейшее размножение тли, осуществляемое партеногенетически, происходит уже на корнях двудольных травянистых растений, на которых может развиваться несколько поколений бескрылых партеногенетических переселенцев. Осенью в потомстве переселенцев могут появляться крылатые самки-полоноски, перелетающие обратно на кусты фисташки, где они рождают самцов и самок, после спаривания откладывающих оплодотворенные яйца в трещины коры. Однако на корнях травянистых растений бескрылые партеногенетические самки могут и зазимовать, и следующей весной продолжать размножаться, рождая живых личинок. Переселенцы белой корневой тли очень многоядны. Известно около 100 видов различных травянистых двудольных, на корнях которых могут размножаться, иногда в массе, эти тли.
Белая корневая тля в отдельные годы сильно вредит табаку, свекле, фасоли и другим культурным растениям, причем нередко в таких местностях, как лесостепная полоса Украины, где фисташки нет. В этих случаях вид существует только в одной форме — бескрылых переселенцев. Такая форма тли, утратившая связь с основным кормовым растением, называется неполноциклой, поскольку выпадают обоеполое и крылатые поколения. Расселению тли в таких случаях, правда на недалекие расстояния, способствуют посещающие их муравьи, слизывающие сладкие экскременты тлей.
Встречаемость белой корневой и других видов тлей, исходно связанных с фисташкой, на корнях травянистых растений в местностях, где фисташка не растет и расти не может, наш известный энтомолог А. К. Мордвилко объяснил тем, что в доледниковый период ареал фисташки соответствовал современной области распространения бескрылых корневых тлей этого и близких видов (триба Fordea). Специалисты по исторической географии растений сперва недоверчиво отнеслись к предположению специалиста по тлям, убежденно отстаивавшего свою теорию. Тем большим был триумф А. К. Мордвилко, когда распространение фисташки в доледниковый период было бесспорно доказано методом пыльцевого анализа: в доледниковых отложениях в местностях, где теперь распространена белая корневая тля, была обнаружена пыльца фисташки!
Поскольку белая корневая тля в отдельные годы причиняет серьезные повреждения посевам разных культур в тех районах, где этот вредитель встречается только в неполноциклой корневой форме, неспособной к расселению на большие расстояния, посевы двудольных культур следует чередовать с посевами зерновых, на корнях которых эта тля питаться не может. Такое простое агротехническое мероприятие предохраняет ценные технические культуры от повреждений белой корневой тли.
Тли — очень богатая видами группа насекомых. Очень быстрое размножение, связанное с небольшой продолжительностью развития, способностью к партеногенезу, с живорождением, приводит к быстрым изменениям наследственных особенностей тлей. Отсюда и обилие их видов. В настоящее время описано около 20 000 видов тлей, но их, несомненно, много больше.
У медяниц и тлей много черт регрессивного развития, связанных с большой плодовитостью, легкостью добывания пищи и малой подвижностью. Личинки тлей и бескрылые тли очень мало и неохотно передвигаются. Еще ярче эти черты выражены у червецов и щитовок.
Источник